
как настроить и как он работает?
Контекстная реклама
Автор Сергей Шевченко На чтение 6 мин Просмотров 190 Опубликовано
Содержание
- Когда необходим временной таргетинг?
- В каких сферах эффективен временной таргетинг?
- Как работает временной таргетинг в контекстной рекламе?
- Как настроить временной таргетинг в Google Ads?
- Что учитывать при настройке временного таргетинга?
Временной таргетинг используется для ограничения показов объявлений в определённые периоды времени. Границы этих промежутков задаёт рекламодатель. Механизм обычно используют в целях минимизации неэффективного расходования рекламного бюджета.
Актуальность применения временного таргетинга напрямую связана с колебанием спроса на продукт в течение суток, недели или какого-либо иного заданного периода времени.
Любой рекламный сервис в онлайн-пространстве позволяет настраивать ограничения по времени. Такой таргетинг уменьшает количество непродуктивных показов объявлений — кампания останавливается в те моменты, когда реклама не может оказывать воздействие на целевую аудиторию. Регулируются показы в зависимости от поры года, месяца, выходных и рабочих дней, дней недели и времени суток.
Когда необходим временной таргетинг?
Существует несколько ситуаций, в которых применение временного таргетинга в контекстной рекламе оправдано и эффективно. Рассмотрим наиболее популярные из них:
- Нужно отключить рекламу в нерабочее время. Это относится, например, к организациям, оказывающим услуги по офисному графику. Привлечённый с помощью платного объявления посетитель может оставить заявку сразу после ознакомления с сайтом.
 Таргетинг рекламы по времени сильно увеличивает шанс на конверсию такого пользователя в реального клиента. В случае, когда кампания не останавливается все 24 часа в сутки, а компания работает с 10 до 19, следует разместить на сайте форму обратной связи. Это уменьшит риск потери клиента;
Таргетинг рекламы по времени сильно увеличивает шанс на конверсию такого пользователя в реального клиента. В случае, когда кампания не останавливается все 24 часа в сутки, а компания работает с 10 до 19, следует разместить на сайте форму обратной связи. Это уменьшит риск потери клиента; - Требуется продвинуть сезонные акции и предложения. При ярко выраженном сезонном спросе часто необходимо запустить рекламу и увеличить ставки именно в праздничные или выходные дни;
- Невозможно воздействовать на ЦА в определенное время суток. Например, бессмысленно заниматься продвижением детских товаров после полуночи, а показы объявлений об услугах для 30-40-летней аудитории лучше сократить в дневные часы, ведь в это время большинство потенциальных клиентов находится на работе;
- Предложение продукта ограничено по времени. Применимо для акции, срок действия которой заканчивается, например, 30 сентября. Если организация при этом работает до 20:00, то остановить рекламную кампанию следует уже в 19:00. Ведь те, кто ознакомится с предложением, должны ещё успеть добраться до магазина, чтобы купить акционный продукт;
- Клиенты компании находятся в разных часовых поясах. В такой ситуации рекомендуется сдвигать показы с учётом разницы во времени;
- Требуется сократить рекламные затраты. Используя инструменты таргетинга, Вы можете сэкономить бюджет, не снижая эффективности кампании.
 Таргетинг рекламы по времени сильно увеличивает шанс на конверсию такого пользователя в реального клиента. В случае, когда кампания не останавливается все 24 часа в сутки, а компания работает с 10 до 19, следует разместить на сайте форму обратной связи. Это уменьшит риск потери клиента;
Таргетинг рекламы по времени сильно увеличивает шанс на конверсию такого пользователя в реального клиента. В случае, когда кампания не останавливается все 24 часа в сутки, а компания работает с 10 до 19, следует разместить на сайте форму обратной связи. Это уменьшит риск потери клиента;/f/86518/3152x2027/67836cb8c1/shutterstock_290015303.jpg) Ведь те, кто ознакомится с предложением, должны ещё успеть добраться до магазина, чтобы купить акционный продукт;
Ведь те, кто ознакомится с предложением, должны ещё успеть добраться до магазина, чтобы купить акционный продукт;Совет! Перед тем, как настроить таргетинг по времени, следует тщательно проанализировать рынок. Чем точнее Вы определите периоды онлайн-активности целевой аудитории, тем успешнее будет Ваша реклама.
В каких сферах эффективен временной таргетинг?
Рассмотрим наиболее распространённые сферы, где кампании с временным таргетингом работают гораздо эффективнее непрерывной рекламы:
- Услуги такси, няни. Пиковая нагрузка приходится на вечернее время в пятницу, на выходные и праздничные дни;
- Доставка готовых блюд. Наиболее активна целевая аудитория вечером;
- Билеты на мероприятия, концерты, спектакли и в кино чаще покупают после обеда — около 14:00. Второй пик начинается в 18 и заканчивается в 20 часов;
- Любые предложения, имеющие отношение к офисной деятельности, правильнее продвигать в стандартные рабочие часы — в период с 9 до 17.
 Наиболее активна целевая аудитория вечером;
Наиболее активна целевая аудитория вечером;Важно! Применение временного таргетинга не исключает возможность показа рекламы в другие интервалы. Всегда следует проверять вероятность привлечения большего числа клиентов в «выгодное» время при размещении на более высоких позициях.
Чтобы определить оптимальный интервал показа рекламы, анализируйте статистические данные и экспериментируйте. Например, можно запустить две идентичные кампании до и после полудня и сопоставить стоимость перехода и привлечения клиента спустя какое-то время.
Как работает временной таргетинг в контекстной рекламе?
В системе Google Ads всем рекламодателям доступны гибкие настройки временных ограничений для показа объявлений. Чтобы понять, как работает временной таргетинг, рассмотрим наиболее простые схемы:
Чтобы понять, как работает временной таргетинг, рассмотрим наиболее простые схемы:
- В соответствии с заданным рабочим режимом. Пользователи видят рекламу только в рабочие часы организации;
- Показ в круглосуточном режиме. Эта стратегия обычно обходится довольно дорого и применима только в случае, когда сама организация работает 24/7 или на её сайте есть форма заказа или обратной связи;
- Запуск кампании только в рабочие дни. Объявления демонстрируются без перерыва пять дней в неделю и не показываются в выходные;
- Только в субботу и воскресенье. Стратегия актуальна для бизнесов, спрос на услуги которых появляется или значительно увеличивается только в выходные дни;
- Первая или вторая половина дня. Показы привязываются к интервалу, когда целевая аудитория наиболее чувствительна к рекламе конкретного продукта;
- Сезонный таргетинг. Если товары и услуги чётко ориентированы на какой-либо праздник или время года, то такая стратегия является оптимальной.

Как настроить временной таргетинг в Google Ads?
Чтобы настроить временной таргетинг в Google Ads, на странице соответствующей кампании в разделе «Расписание показа объявлений» кликните на «Изменить расписание показа объявлений». Возможность ограничить показы объявлений по времени есть только в кампаниях подтипа «Все функции»:
В появившемся модуле, выбирая требуемые дни и время, Вы можете установить или скорректировать режим показа рекламы.
Помимо этого, система позволяет задать корректировку ставок по времени показа рекламы. Чтобы их настроить, следует отметить соответствующие позиции и кликнуть на «Изменить корректировки ставок». Эта функция не работает в кампаниях, где, в соответствии с выбранной рекламодателем стратегией, ставки назначаются автоматически.
В открывшемся блоке укажите значение корректировки в процентном отношении и выберите «Уменьшить»/«Понизить».
Что учитывать при настройке временного таргетинга?
- Настроить корректировку ставок можно только для стратегий, управляемых вручную.
- При настройке временного таргетинга важно не забывать о разнице часовых поясов. Если показ объявлений необходимо ограничить по времени, а целевая аудитория находится в нескольких регионах, то для каждой часовой зоны запускайте отдельную кампанию;
- Ограничение демонстрации рекламы в некоторые часы всегда приводит уменьшению трафика. Иногда такие потери могут быть значительными. Прежде чем изменить настройки кампании, подумайте над тем, зачем Вы это делаете и какие результаты ожидаете получить. Вполне может оказаться, что тотально ограничивать показы в определённые интервалы времени не следует, достаточно лишь понизить ставку на этот период. Такой «мягкий таргетинг» помогает не только сохранить трафик, но и сэкономить бюджет.

Оцените автора
Сергей Шевченко Google-логистПакет услуг «90 дней Google рекламы», поможет сделать Вашу рекламную кампанию не только выгодной, а и увеличить продажи с нее
ДЕТАЛЬНЕЕ
Как работает временной таргетинг в Яндекс Директ
Введение
Временной таргетинг – это один из инструментов таргетированной реклпмы( Google Ads, Facebook Ads, Яндекс Директ) который помогает повысить эффективность рекламного объявления и не тратить рекламный бюджет в никуда. Зачем он нужен, и как его использовать, сейчас мы подробно всё это разберем.
Зачем он нужен, и как его использовать, сейчас мы подробно всё это разберем.
Я расскажу вам, зачем нужен временной таргетинг, рассмотрим подводные камни этого инструмента, а также разберем различия между временным таргетингом в Google Ads и Яндекс Директ, есть ли существенные различия между этими рекламными сервисами.
Поговорим о том, как настраивать временной таргетинг и я поделюсь с вами маленькими секретами в настройки временного таргетинга.
Зачем нужен временной таргетинг?
Зачем и кому нужен временной таргетинг, как им пользоваться? Каждая настройка таргетированной рекламы разделяется на несколько этапов, в каждой социальной сети эти этапы различаются. Но, именно этот инструмент можно использовать везде. А вот использовать или нет это уже решать заказчику и исполнителю.
Сейчас мы углубимся в Google Ads и Яндекс Директ. Так как здесь этот инструмент используется чаще всего.
Что необходимо учитывать настраивая временный таргетинг?
Временной таргетинг полезная функция, которая используется для привлечения клиентов в определенный период времени, таким образом, можно показывать рекламные объявления в определенный период времени.
Например: Если у вашего клиента сервис такси по вызову, то вам стоит крутить рекламные объявления целый день, потому что люди пользуются услугами такси постоянно. А вот если у вас бесплатная доставка еды до обеда, то рекламу стоит показывать с 00:00 до 12:00, именно в это время люди смогут заказать у вас еды, или если они увидят рекламу ночью или утром, то смогут успеть заказать еду бесплатно.
И прекращать показ рекламы.
Но, каждому рекламодателю необходимо понимать, что их клиенты могут увидеть рекламное объявление утром, а сделать заказ только через несколько дней. Поэтому не стоит надеяться только на временной таргетинг, а лучше использовать его в связке с рекламной компанией без временного таргетинга.
Также, стоит учитывают целевую аудиторию, на которую настраивается реклама. Чаще всего нужно обращать внимание на рабочее время людей, это можно определить логически.
Например: если ваша целевая аудитория студенты, то скорее всего временной таргетинг нужно ставить на вечернее и ночное время, так как утром и днём студенты на учебё или выполняют задания связанные с учёбой.
Как работает временной таргетинг в Яндекс Директ и Google AdWords?
Принципы работы временного таргетинга в Яндекс Директ и Google AdWords практически идентичны. Поэтому 95% написанной информации относится к обоим рекламным сервисам.
По умолчанию в каждом рекламном кабинете временной таргетинг установлен на показ рекламных объявлений круглосуточно, то есть 24 часа без остановки.
Этот инструмент позволяет выбрать дни и определенные часы для показа рекламных объявлений нужной аудитории.
Всё-таки зачем нужно использовать временной таргетинг?
Ответ довольно прост: Чтобы увеличить или уменьшить количество кликов по рекламному объявлению в определенные дни/часы, а также уменьшить стоимость результата (клик, переход на страницы, лид). А также временной таргетинг отлично подойдёт для крупных бизнесов, которые занимаются сезонными товарами.
Ведь можно выделить сезонные предложения и акции. С этой целью нужно, наоборот, запустить объявления и поднять ставки именно в выходные дни или праздники. Так как большинство людей совершают крупные покупки или покупают подарки именно на выходных
Так как большинство людей совершают крупные покупки или покупают подарки именно на выходных
Мы поняли, как работает и зачем нужен такой инструмент как «временной таргетинг», а сейчас мы разберем отдельно временной таргетинг в Яндекс Директ и Google AdWords и сравним этот инструмент в двух этих рекламных сервисах.
Яндекс Директ
В Яндекс Директ временной таргетинг практически ничем не отличается от других рекламных сервисов.
Безусловно интерфейс у всех разный, но суть использования инструмента «одна и та же.
Иногда не стоит ограничивать показ рекламных объявлений временным таргетингом, так как в таких бизнесах как кафе, коворкинг, химчистка и тд. Люди могут увидеть рекламу в любое время дня и сохранив контакты обратиться за услугой позже. Но, реклама с временным таргетингом должна различаться содержанием в рабочее и нерабочее время вашей компании.
Например: В рабочее время вы можете запустить рекламу с призывом к действию: написать, позвонить, купить что-либо.
В Яндекс Директ временной таргетинг настраивается следующим образом:
В блоке расписание показов на странице параметров установите время когда вы хотите показывать когда вы хотите показывать свою рекламу.
Как это сделать?
Есть два способа:
-
Использовать имеющиеся в рекламном кабинете Яндекс Директ шаблоны.
-
Сделать это в ручную, отметив в таблице нужные вам дни и часы.
Также в Яндекс Директ общее количество часов показа рекламы не должно быть меньше 40 часов в неделю.
Также кроме дней и часов необходимо выбрать страну и часовой пояс. В Яндекс Директ для одной рекламной компании выбрать только один часовой пояс и если вам нужно настроить рекламу на города с разными часовыми поясами, то лучше всего создать отдельную рекламную компанию или рекламное объявление для каждого города.
Временной таргетинг в Яндекс Директ позволяет повышать ставки от 0% до 200%.
Когда мы сделаем ставку 0% то реклама остановится и перестанет показываться. А если повысить ставку до 200%, то рекламный бюджет увеличится в два раза.
Google AdWords
Временной таргетинг в Google AdWords идентичен с Яндекс Директ и настроить этот инструмент можно на уровне компании рекламных объявлений. В Google AdWords рекламодатель может выбрать два варианта насктройки временного таргетингв:
-
Отдельно будние дни и отдельно выходные
-
Выбрать отдельные дни
Сейчас я покажу короткую инструкцию по настройке такого инструмента рекламы как «временной таргетинг» в Google AdWords:
-
Естественно что в первую очередь нам нужно определится с рекламной компанией и выбрать её.
-
Открыть страницу «Расписание показов объявления»
-
Далее нужно открыть столбец «Корректировка ставок»
-
Для того чтобы открыть окно с настройками нужно нажать на нужное число в столбце «Корректировка ставок».

Инструмент «Временной таргетинг» в Google AdWords позволяет понижать и повышать ставку в процентах от 90% до 900%. Вот вам и существенная разница с Яндекс Директ.
В Яндекс Директ этот показатель варьируется от 0% до 200%.
Советы по настройке временного таргетинга
-
Корректировка ставок доступна только для стратегий с ручным управлением ставками. Как в Яндекс.Директе, так и в Google AdWords при смене стратегии на автоматическую настройки не сохраняются.
-
Как я и писал ранее важно учитывать часовой пояс. Ведь если вы ограничиваете временной таргетинг, то для регионов с разными часовыми поясами нужно создать отдельные кампании.
-
С помощью инструментов аналитики изучайте аудиторию, тестируйте показ в разное время. Эти результаты помогут выделить целевую аудиторию, найти постоянных клиентов и улучшить эффективность рекламы.
Как известно всё познаётся в сравнении:
Для сравнения возьмём Facebook Ads. В отличии от Яндекс Директ и Google AdWords временной таргетинг в Facebook Ads используется не так часто и не приносит столько результата сколько в контекстной рекламе. В Facebook Ads делается упор на количество показов рекламы как можно большему количеству потенциальных клиентов, в то время как в контекстной рекламе часто используется временной таргетинг как один из основных инструментов.
В отличии от Яндекс Директ и Google AdWords временной таргетинг в Facebook Ads используется не так часто и не приносит столько результата сколько в контекстной рекламе. В Facebook Ads делается упор на количество показов рекламы как можно большему количеству потенциальных клиентов, в то время как в контекстной рекламе часто используется временной таргетинг как один из основных инструментов.
Пример
Для правильного использования временного тагетинга необходимо в первую очередь определить целевую аудиторию, это нужно чтобы понять в какое время клиенты будут чаще всего делать заказы на доставку еды. И сделать акцию чтобы стимулировать людей заказывать именно у этой компании, а не у их конкурентов. Тут временной таргетинг поможет нам переманить больше клиентов на свою сторону, ведь даже когда кончится действие акции, если сервис отвечают стандартам качества то эти клиенты также останутся с этой компанией
В качестве примера можно разобрать использование инструмента «Временной таргетинг» в рекламной компании Донерной, которая осуществляет доставку бесплатно с 10:00 до 14:00. Почему именно это время?
Почему именно это время?
Первый вариант: В это время мало продаж и сделав доставку бесплатной рекламодатель хочет привлечь внимание.
Второй вариант противоположен первому, в этом случае рекламодатель наоборот поощряет своих клиентов, так как в это время большое количество заказов.
Здесь нельзя 100% сказать что правильно, а что нет. Также нельзя сказать будет ли временной таргетинг улучшать результат или нет. Всё нужно тестировать, пробовать и только после тестов находить работающие компании и масштабировать их.
Заключение
В заключение ко всей выше перечисленной информации могу добавить своё личное мнение касательно такого инструмента как временной таргетинг. Этот инструмент есть во всех рекламных кабинетах и он должен тестироваться в каждой рекламной компании так как таргетированная реклама это тесты и никто не может с 100%; уверенностью сказать что это будет работать, а вот это не будет работать с определенной нишей, так как есть опыт когда не работало. Таргетированная реклама интересна тем, что в ней два абсолютно идентичных рекламных группу с вероятностью в 80% принесут разные результаты.
Таргетированная реклама интересна тем, что в ней два абсолютно идентичных рекламных группу с вероятностью в 80% принесут разные результаты.
Например есть колоссальное отличие между контекстной рекламой(Google AdWords и Яндекс Дзен) и таргетированной рекламой(Facebook, VKontakte, Tik-Tok) в использовании такого инструмента как «временной таргетинг». И это обуславливается рядом факторов, один из них это то что в Facebook Ads таргетологи крутят рекламу 24 часа подряд, чтобы не навредить рекламной компании и не сделать цену за результат дороже. Безусловно, мы используем временной таргетинг, но не так часто и он не играет особой роли в таргетированной рекламе Facebook Ads.
Эти сервисы помогут в ваших начинаниях:
| ВК Сёрфинг | Tooligram Media |
| Supa | SMMbot |
Если вы обнаружили ошибку в тексте, выделите её и нажмите ctrl + enter
Как работает временной таргетинг в Яндекс.
 Директе и Google AdWords
Директе и Google AdWordsНаталия Корсунова
Маркетолог-аналитик
5 минут
6362
4
Один из наиболее важных факторов для настройки контекстной рекламы в Яндекс.Директе и Google AdWords — правильный временной таргетинг. Но как понять, когда пользователь максимально расположен к совершению заказа? Какие факторы следует учесть при выборе времени показа объявлений? Попробуем рассмотреть основные механизмы подбора оптимального расписания работы рекламы.
Временной таргетинг всегда должен основываться на особенностях целевой аудитории рекламодателя. Также важными факторами являются сезонность и механизм продаж в конкретной тематике.
Как работает эта схема?
- Рекламодатель должен составить портрет потенциального покупателя. Следует учесть пол и возраст, занятость, материальное положение, попробовать представить, как клиент проводит день и когда он максимально активен в интернете. Например, если Ваш потенциальный покупатель — это москвич, работник офиса, активно использующий смартфон, то наиболее «горячее» время для показа рекламы такому пользователю — с 17:00 до 20:00. Это час пик, когда большое количество людей едет домой после работы и использует смартфон для поиска товаров и услуг.
- К сформированному представлению о целевой аудитории следует добавить особенности механизма продаж для конкретного товара. К примеру, пользователь более активно будет искать место, куда можно съездить отдохнуть на выходные, в четверг и пятницу. Не стоит забывать и об отложенном спросе. Если тематика сайта предполагает большое время принятия решения, нет смысла выставлять жесткие ограничения временного таргетинга. Пользователь может изучать предложения в любое время суток и в любой день недели.
- Важно учесть время работы офиса рекламодателя. Если связаться с рекламодателем можно только в рабочее время по будням, то стоит включать объявления только в это время. Необработанные в короткие сроки заявки и звонки не принесут прибыли.
- Не стоит забывать и о часовых поясах. Рабочий день за Уралом начинается намного раньше, чем в Москве. Если большой поток заказов поступает из восточных регионов, стоит настроить старт рекламы с учетом часового пояса клиентов и предложить пользователям форму обратного звонка с выбором удобного времени для связи.
- Все тематики в контекстной рекламе зависят от сезонности, выходных и праздничных дней. В выходные спрос в тематиках B2B существенно падает. Это можно объяснить тем, что зачастую услугами и товарами для B2B интересуется не собственник бизнеса, а сотрудник компании, который проводит поиск только в рабочее время. Например, не стоит транслировать рекламу товаров для офиса в субботу и воскресенье. Тем не менее, по выходным дням резко возрастает активность пользователей в социальных сетях, поэтому тематики с низкой стоимостью товаров, актуальные для соцсетей, могут получить больше кликов и конверсий.
 Это час пик, когда большое количество людей едет домой после работы и использует смартфон для поиска товаров и услуг.
Это час пик, когда большое количество людей едет домой после работы и использует смартфон для поиска товаров и услуг. Если большой поток заказов поступает из восточных регионов, стоит настроить старт рекламы с учетом часового пояса клиентов и предложить пользователям форму обратного звонка с выбором удобного времени для связи.
Если большой поток заказов поступает из восточных регионов, стоит настроить старт рекламы с учетом часового пояса клиентов и предложить пользователям форму обратного звонка с выбором удобного времени для связи.При любых изменениях временного таргетинга следует отслеживать изменения конверсии. В некоторых тематиках верное время показа объявлений является основным фактором достижения конверсии. Наверное, наиболее ярко зависимость процента конверсии от расписания работы рекламы можно проследить в тематике доставки еды.
Наверное, наиболее ярко зависимость процента конверсии от расписания работы рекламы можно проследить в тематике доставки еды.
К примеру, возьмем рекламодателя, который занимается доставкой осетинских пирогов и блюд восточной кухни. После анализа целевой аудитории и серии тестирований было установлено, что наиболее конверсионными днями недели для рекламы являются понедельник и пятница. Именно в эти дни пользователи больше всего нацелены на совершение покупки. Наиболее конверсионным временем суток является период с 9:00 до 15:00.
Как работает временной таргетинг в Яндекс.Директе?
- В блоке «Расписание показов» укажите страну и корректный часовой пояс, выбрав из предложенного списка. По умолчанию стоит Москва.
- Яндекс.Директ автоматически переходит на летнее и зимнее время.
- Можно выбрать только те часовые пояса, которые отличаются от времени по Гринвичу на полное количество часов (на 2 или 3, но не на три с половиной).
- Параметры рекламы не зависят от выбранного часового пояса. Начало кампании и данные отчетов статистики всегда показываются по Москве.
- Временной таргетинг в Яндекс.Директе позволяет выбрать только один часовой пояс. Если Вы рекламируете продукт в городах с другим временем, создавайте для них отдельные рекламные кампании.
- Временной таргетинг в Директе позволяет изменить показ рекламы в выходные и праздники. Учитываются только официальные государственные праздники.
- Поставьте галочку в пункте «Учитывать праздничные дни».
- Укажите промежуток времени.
- Поставьте галочку в пункте «Учитывать рабочие выходные», если рабочий день перенесен на субботу или воскресенье. Тогда пользователи будут видеть объявление так же, как в будни.
Таким образом, в выходные и праздники можно показывать рекламу в определенные часы или вообще не демонстрировать ее.
- Временной таргетинг позволяет корректировать ставки. В определенное время показа программа может автоматически повышать и понижать стоимость клика.
- Включите режим управления ставками.
- Установите размер. На сколько изменится цена в процентах от 0 до 200, где 100% — текущая ставка, а 0% — отсутствие показов.
- Укажите время, когда ставка должна увеличиться или уменьшиться.
Настройка работает только при ручном управлении ставками и сбрасывается при переходе на автоматическое.

 В определенное время показа программа может автоматически повышать и понижать стоимость клика.
В определенное время показа программа может автоматически повышать и понижать стоимость клика.
Как работает временной таргетинг в AdWords?
- По умолчанию программа демонстрирует рекламные объявления каждый день 24 часа в сутки. С помощью временного таргетинга в AdWords можно выбрать дни показа и время.
- Нажмите «Расписание показа объявлений» в настройках.
- Выберите «Создать особое расписание».
- Укажите дни и часы, когда реклама будет работать.
Время соответствует часовому поясу в аккаунте.
Вы можете изменить его только раз через справочную Google AdWords. Составляя расписание показов, учитывайте часовой пояс клиентов из других городов. Нельзя выбрать временной период, который охватывает 2 дня — например, с 18:00 до 02:00. В программе новый день начинается в полночь. Поэтому нужно выставить 2 временных периода: с 00:00 до 02:00 и с 18:00 до 00:00. - Нажмите «Сохранить».
Чтобы проверить настройки таргетинга, зайдите в настройки → расписание показов объявлений → сведения о расписании → часы и дни недели.
- В AdWords можно увеличить или уменьшить стоимость клика в разные часы, изменив расписание показа объявлений. Например, если Ваша целевая аудитория неактивна в обеденный перерыв, стоит понизить ставки с 12:00 до 13:00. С помощью временного таргетинга показ уменьшится, и Вы не будете зря расходовать рекламный бюджет.
 Вы можете изменить его только раз через справочную Google AdWords. Составляя расписание показов, учитывайте часовой пояс клиентов из других городов. Нельзя выбрать временной период, который охватывает 2 дня — например, с 18:00 до 02:00. В программе новый день начинается в полночь. Поэтому нужно выставить 2 временных периода: с 00:00 до 02:00 и с 18:00 до 00:00.
Вы можете изменить его только раз через справочную Google AdWords. Составляя расписание показов, учитывайте часовой пояс клиентов из других городов. Нельзя выбрать временной период, который охватывает 2 дня — например, с 18:00 до 02:00. В программе новый день начинается в полночь. Поэтому нужно выставить 2 временных периода: с 00:00 до 02:00 и с 18:00 до 00:00.- Предыдущий материал
- Следующий материал
что это такое, как его настроить в Яндекс Директе
Получи нашу книгу «Контент-маркетинг в социальных сетях: Как засесть в голову подписчиков и влюбить их в свой бренд».
Подпишись на рассылку и получи книгу в подарок!
Временной таргетинг — это специальный рекламный механизм, с помощью которого можно выбрать время показа объявлений.
Чтобы понять, что такое временной таргетинг, рассмотрим простую аналогию. Всем известно, что в разное время по телевизору показывают разную рекламу. Рекламодатели знают, что реклама игрушек в ночное время или реклама мужских товаров во время мыльных опер будет иметь минимальный эффект, так как целевая аудитория не увидит объявление. Все мы знаем, что размещение рекламы в прайм-тайм стоит намного дороже, так как охват аудитории шире. Из этого следует, что при размещении рекламы важно учитывать время ее показа. В противном случае вы рискуете потратить рекламный бюджет впустую. Временной таргетинг позволяет четко определить время показов рекламы в интернете.
Зачем нужен временной таргетинг
Существует несколько важных причин, почему его нужно использовать:
- Исключить показы рекламы в нерабочее время. У вас выходной, а пользователь увидел объявление, перешел по ссылке, но не смог дозвониться до вас.
- Привлечь внимание к сезонным товарам и акциям.
- Работать с целевой аудиторией.
- Чтобы увеличить эффективность кампании. Если кампания активна, когда активна ЦА, эффект сразу же увеличивается.
- Сократить расходы. Не показывать объявление, когда целевая аудитория спит. Расходы ниже, конверсия выше.
 У вас выходной, а пользователь увидел объявление, перешел по ссылке, но не смог дозвониться до вас.
У вас выходной, а пользователь увидел объявление, перешел по ссылке, но не смог дозвониться до вас.Как работает временной таргетинг в Яндекс Директ и Google AdWords
Временной таргетинг Яндекс и Гугл работает согласно выбранному вами режиму. То есть вы устанавливаете, когда будут отображаться объявления. Временной таргетинг работает по следующим схемам:
- Во время работы компании. Клиент видит объявление только тогда, когда компания работает, то есть когда у него есть возможность позвонить/сделать заказ/получить консультацию и т.д.
- На протяжении всей недели. Объявления будут показываться 7 дней в неделю, 24 часа в сутки. То есть, объявления показываются непрерывно. Данный тариф подойдет практически для любого бизнеса, однако и стоит он не дешево.
- 5 дней в неделю. Реклама показывается только по будням, с понедельника по пятницу.
- В выходные. Здесь наоборот, предложения транслируются в субботу и воскресенье.
- В сезон/определенный период. Рекламные объявления показываются только летом/зимой/в преддверии праздников и т.д.
 Данный тариф подойдет практически для любого бизнеса, однако и стоит он не дешево.
Данный тариф подойдет практически для любого бизнеса, однако и стоит он не дешево.Яндекс Директ
Начнем с того, как выглядит временной таргетинг Директ. Как вы видите, здесь много различных опций для выбора. На данном рисунке выбрана «Пятидневка», объявления отображаются только в рабочее время.
Обратите внимание на опцию «Часовые пояса». Здесь можно выбрать часовой пояс для города, где находится ваша компания, будь то Магадан, Владивосток или Калининград. Кроме Российской Федерации, сервис позволяет выбрать часовой пояс и для стран-участниц СНГ. Помимо этого, часовой пояс можно установить для различных других стран мира, на выбор представлена 231 страна.
Еще одна полезная опция, которую нужно настроить в Яндекс Директ — режим управления ставками. Здесь можно установить стоимость клика как на основное время показа, так и на оставшиеся временные периоды (здесь устанавливается часть от стандартной стоимости клика). За счет этого удается увеличить приток клиентов при том же уровне расходов, а также повысить объем показов.
Здесь можно установить стоимость клика как на основное время показа, так и на оставшиеся временные периоды (здесь устанавливается часть от стандартной стоимости клика). За счет этого удается увеличить приток клиентов при том же уровне расходов, а также повысить объем показов.
Как вы видите на рисунке, 100% стоимости приходится на первоначально установленную «Пятидневку» в рабочее время, 50% — на клики в эту еже пятидневку, но нерабочее вечернее время и 20% — на выходные.
Следующая опция —учет рабочих дней. Например, у вас настроен показ только в рабочие дни. Если праздник выпадет на рабочий день, объявления не будут. Обратите внимание, что здесь учитываются только выходные и праздники Российской Федерации.
Другой механизм — учет праздничных дней. С его помощью система включает/отключает показы при необходимости. Также здесь можно настроить другой режим показов объявлений во время праздников. В сочетании с опцией режима управления ставками, здесь можно установить часть от стоимости клика для показов в праздничные дни. Здесь алгоритмы также берут во внимание только праздники РФ.
Здесь алгоритмы также берут во внимание только праздники РФ.
Google AdWords
Система Гугл также предлагает своим пользователям воспользоваться временным таргетингом.
Осуществляется это в три этапа:
- Часовой пояс указывается во время регистрации аккаунта на сервисе Google AdWords.
- В расширенных настройках кампании нужно выбрать дату ее начала и окончания, спланировать показ рекламных объявлений.
- Настроить основное расписание показа.
После этого, по желанию уже настраиваются дополнительные возможности.
Часто используется установка корректировки ставок. Можно скорректировать цену за клик в определенные дни, выделить один/несколько дней и в течение выбранного периода повысить/понизить стоимость за клик.
Популярностью пользуется и выбор метода показа. Их всего два: ускоренный и стандартный. При стандартном показы распределяются равномерно согласно выбранному графику. То есть реклама показывается на протяжении выбранного времени, но не при каждом запросе пользователей.
Основная цель — растянуть показы на весь день, а не израсходовать все в первые несколько часов. При ускоренном методе, частота показов объявлений повышается, пока зафиксированный бюджет не израсходуется полностью. То есть, при каждом запросе пользователя (если в нем есть ваши ключевые слова), будет отображаться объявление.
Как вы видите, временной таргетинг позволяет повысить эффективность рекламных объявлений, а также оптимизировать расходы.
TrueDigital — говорим начистоту о digital
|
Artics Internet Solutions
Подготовили очередную подборку кейсов для вдохновения. На этот раз поговорим про то:
Настройка O2O-атрибуции во время масштабных кампаний к «Черной пятнице» и Новому году позволила отследить более 30 тыс продаж в офлайн-точках по итогу онлайн-кампаний. В чем суть: основная доля продаж ювелирной сети «585*Золотой» приходится на офлайн-магазины. При этом во время сезонных акций бренд проводит масштабные онлайн-кампании, и чтобы оценить их эффективность и оптимизировать кампании сети, команда настроила О2О-атрибуцию. В результате бренд выяснил, что пользователи совершили более 30 000 покупок в офлайн-магазинах после взаимодействия с рекламными объявлениями в интернете, а ДРР составил 3%. Как это работает: в качестве одной из площадок для запуска флайта был выбран myTarget. Таргетировались на новую аудиторию (по интересам, конкурентам, lool-alike) и на существующих, так называемых «заснувших» пользователей, которые когда-то покупали у бренда, но делали это давно. Отслеживание покупок в магазинах после старта кампании происходило через инструмент «Офлайн-конверсии». На площадке настроили опцию для атрибуции визитов пользователей, которые видели рекламные объявления кампании. В качестве источника данных об офлайн-покупках использовали данные CRM-базы «585*Золотой» (телефоны + сумма заказа). Поскольку площадка при загрузке данных хеширует их, подобрали ключ и уже после этого настроили учет конверсий. В счет брали только последнее взаимодействие с рекламой. В результате увидели, что некоторые кампании, приносившие по 5-10 онлайн-заказов, давали более 200 офлайн-заказов. Почему это работает: офлайн-трекинг и ориентир на последнее взаимодействие с рекламой помогли не только оценить, насколько кампания эффективна в рамках продаж, но и вовремя оптимизировать кампании и найти объявления, которые приносили больше конверсий. Стратегия кампании была подобрана так, чтобы проработать максимальное количество аудиторий по разным таргетингам — это помогло выйти на пользователей, которые наверняка совершат покупку. Если говорить про O2O-атрибуцию, то была проделана большая работа — мы отформатировали данные миллионов пользователей, чтобы понять, насколько digital-реклама была эффективной. Также мы увидели, какие аудитории и таргетинги сработали лучше всего. Мария Самойлова, специалист по таргетированной рекламе Artics Internet Solutions |



|
«Додо Пицца»: временный таргетинг и меню к завтраку, обеду и ужину
Использование временного таргетинга позволило показать потребителю релевантное объявление с меню в зависимости от времени суток — для завтрака, обеда или ужина, — и достичь снижения CPA на 41%. В чем суть: команда сети пиццерий «Додо Пицца» выстроила гипотезу, что пользователи заказывают доставку на завтрак, обед или ужин. В рамках одного из запусков протестировали временной таргетинг: запустили тематические баннеры с предложениями по меню для разного времени суток в Instagram Stories. Тестовые группы имели положительную динамику и сработали эффективнее других групп. Как это работает: команда performance-маркетинга провела анализ меню «Додо Пиццы» и разделила его на предложения для обеда, завтрака и ужина. Для каждого сделали отдельный баннер и разделили объявления по времени показа:
Ориентиром послужил распорядок дня среднестатистического жителя города. Это позволило определить время суток, на которое приходится большее количество покупок по выгодной цене. На Android-устройствах в группе lunch удалось снизить CPA на 41%, а для iOS-устройств лучшие показатели были у группы breakfast, CPA сократился на 34%. Почему это работает: сегментирование по времени позволило выйти к потребителю с персонализированным сообщением и показать предложение, которое его заинтересует. Временной таргетинг особенно эффективен при продвижении услуг доставки готовой еды. Например, человек просыпается утром, хочет позавтракать, открывает ленту в социальной сети и видит релевантное предложение — рекламное сообщение с завтраком, которое его скорее всего заинтересует и заставит совершить конверсию. Евгений Коротаев, руководитель проектов Artics Mobile Solutions |

 Каждому временному промежутку соответствовал персонализированный креатив с уникальным предложением.
Каждому временному промежутку соответствовал персонализированный креатив с уникальным предложением.|
«Петрович»: повышение онлайн-продаж с помощью аналитики больших данных
Данные об офлайн-покупках и онлайн-поведении пользователей позволили выявить интересы потенциальных клиентов, выйти к ним с точным предложением и в результате повысить онлайн-продажи на 15%. В чем суть: во время дачного сезона DIY-ритейлеру «Петровичу» важно было охватить максимальное число новых пользователей и продвигать товары для дома и дачи, доступные в интернет-магазине. Команда проанализировала собственные данные о запросах потребителей и запустила digital-кампанию с таргетингом по интересам и персонализированными товарными предложениями. Для оценки влияния рекламы на реальные покупки провели исследование, которое показало, что товары покупали лучше после взаимодействия с рекламой, а сами онлайн-продажи выросли на 15%. Как это работает: ритейлер объединил данные об офлайн- и онлайн-поведении потенциальных покупателей. Это помогло определить их интересы и разделить аудиторию на сегменты: дачников, тех, кто интересовался строительством или мебелью, изучал статьи о ремонте. Для каждого сегмента подготовили рекламные баннеры и видеоролики с персонализированным товарным предложением и запустили таргетированную рекламу. Для оценки эффективности кампании провели A/B-тестирование — сравнили покупки двух групп пользователей: тех, кто посмотрел рекламу в интернете, и тех, кто ее не видел. Оказалось, что потребители, которые видели рекламное сообщение в интернете, покупают на 15% лучше. Почему это работает: алгоритмы, использованной в продвижении технологии, анализировали интересы пользователей и принимали решение о показе релевантного баннера каждому сегменту аудитории. Вероятно, как и у многих DIY-продавцов, большая часть продаж ритейлера приходилась на физические точки, поэтому сбор информации об офлайн-поведении покупателей помог собрать правдивые и точные данные, которые можно было использовать дальше. При сопоставлении этой информации с онлайн-данными бренд ориентировался не только на данные внутри своей экосистемы — интернет-магазин, взаимодействие с digital-рекламой и т. Это помогает создать и показать персонализированное предложение, а также коммуницировать с аудиторией, которая только похожа на клиента компании, а значит, масштабироваться вне сформированного спроса. Иван Круглов, руководитель проектов Artics Internet Solutions |

 Так, например, те, кто изучал тему строительства, видели предложение о покупке лестницы, плитки или линолеума; тем, кто интересовался мебелью, показывали рекламу дверей, сантехники и т.д.
Так, например, те, кто изучал тему строительства, видели предложение о покупке лестницы, плитки или линолеума; тем, кто интересовался мебелью, показывали рекламу дверей, сантехники и т.д.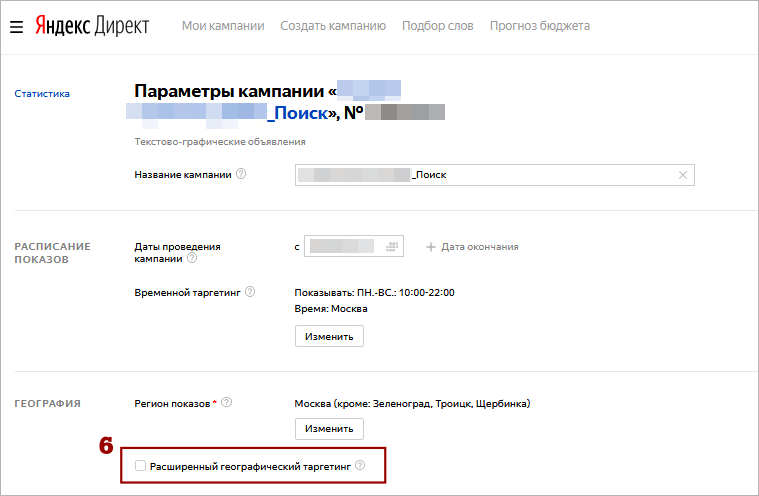 д. Ставку сделали на поиск поведенческих паттернов во вне — какими статьями интересуется человек, хочет ли он сделать ремонт.
д. Ставку сделали на поиск поведенческих паттернов во вне — какими статьями интересуется человек, хочет ли он сделать ремонт. Настройка временного таргетинга
Средний человек озабочен тем, как бы ему убить время, человек же талантливый стремится его использовать.
Артур Шопенгауэр.
Время для показа рекламы — дело очень тонкое и редко когда выставляется верно. Именно к такому умозаключению я пришел, просмотрев тысячи различных аккаунтов и пообщавшись с сотнями клиентов и учеников.
Что же это значит? Более 90% участников рынка пользуются всего 2 стратегиями установки времени для показа рекламы: круглосуточно и время работы офиса. Еще примерно 5% включают показы в низко-конкурентные часы. При первом рассмотрении такие подходы полностью соответствуют всем базовым правилам, как я его называю, «ларькового бизнеса». Однако я вынужден вас разочаровать — более 2/3 всех бизнесов имеют четко выраженное время для показа рекламы, которое не совпадает с классическим представлением.
Еще примерно 5% включают показы в низко-конкурентные часы. При первом рассмотрении такие подходы полностью соответствуют всем базовым правилам, как я его называю, «ларькового бизнеса». Однако я вынужден вас разочаровать — более 2/3 всех бизнесов имеют четко выраженное время для показа рекламы, которое не совпадает с классическим представлением.
Для того, чтобы вы могли избежать таких ошибок в будущем, я предлагаю вам не волшебную таблетку от всех болезней, а несколько примеров и инструментов, которые помогут вам самим научится, как же правильно указывать время для рекламы.
Этап 1. Думаем еще до запуска
Ваша целевая аудитория — живые люди. Более того, это вполне конкретные люди с вполне конкретными привычками, конкретной работой и целым набором потребностей. Для каждой группы целевой аудитории существуют списки схожих потребностей. Я же вам покажу, как с этим можно работать и приведу несколько примеров.
Пример 1. Вы продаете товары для офиса.
Ваша целевая аудитория — b2c сегмент (почему-то принято, что b2b в интернете продавать сложнее b2c, хотя на самом деле ровно наоборот): секретарши, руководители отделов небольших кампаний, ген. директора микробизнеса. Самую большую долю прибыли вам приносят большие клиенты, т.к. это постоянные клиенты с большими заказами, которых, чаще всего, представляют секретарши.
Рис.1 График активности в русскоязычных социальных сетях по данным Cossa. Обратите внимание на время перед обедом и во время обеда.
Секретари обычно выполняют свою работу либо с самого утра (когда получили задачу) с 9 до 11, либо после обеда (что реже), либо перед уходом с работы (чтобы не получить на следующий день). Даже не пытайтесь давать рекламу в обеденный перерыв и перед ним. В обеденный перерыв в принципе бесполезно. А в пред-обеденное время у них и без того много работы. Обратите внимание на график активности в социальных сетях на рисунке 1.
Пример 2. Продаем игровую компьютерную мышь или клавиатуру из дорого сегмента.
Целевая аудитория — офисные работники, студенты из обеспеченной семьи. Для офисных работников самым эффективным временем рекламы будет противоположность предыдущему примеру — они ищут товар для досуга и готовы видеть рекламу в зоне их отдыха и комфорта (социальные сети). Поэтому идеальное время для рекламы будет определятся примерно так:
- обеденный перерыв
- время второго чая 17-00
- а также после 21-00, когда человек расслаблен и отдыхает дома.
Таким образом вы должны разобрать вашу нишу и вашу целевую аудиторию, провести небольшой анализ активности и занятости аудитории и на основании этого сделать вывод — в какое время будет эффективнее всего включать рекламу.
Этап 2. Анализируем конкурентов
Второй шаг, который необходимо предпринять перед запуском, — проверить всех конкурентов и составить сводную таблицу по времени показа. Проделав эту работу, вы сможете сделать сразу несколько предположений:
- Ограничение по времени показа у конкурентов.
- Ограничение по бюджету.
- Наиболее свободное время.

Самые частые стратегии: круглосуточный показ, показ в рабочее время, показ-пока-бюджет-не-кончился. Определяются достаточно просто путем создания таблицы с временными промежутками 1-3 часа (в зависимости от ваших возможностей) и отметкой всех основных конкурентов на предмет (+) есть показ или (-) нет показа.
Рис.2 Анализируем от 3 до 7 конкурентов в течение нескольких дней для выявления благоприятного времени рекламы.
Как только вы закончите эту работу, сразу будет видно, как рекламируется каждый из ваших коллег по рынку. Пример вы можете посмотреть на рис. 2.
Выводов из этой таблицы можно сделать целое множество, многие из которых зависят от вашего знания конкретного рынка и опыта маркетинга в целом. Что необходимо искать, так это наиболее свободное время и наиболее нестандартных конкурентов, которые рекламируются в нестандартное время. Для этого рекомендую использовать более частую шкалу (возможно 1-2 часа). Если показ рекламы конкурента выпадает из какой-либо логичной структуры, например, стабильно с 13 до 15 и с 19 до 00, то, с огромной вероятностью, вы попали на умного конкурента, который подобрал наиболее вкусные часы.
Если показ рекламы конкурента выпадает из какой-либо логичной структуры, например, стабильно с 13 до 15 и с 19 до 00, то, с огромной вероятностью, вы попали на умного конкурента, который подобрал наиболее вкусные часы.
Также если вы знакомы с конкурентами лично и знаете информацию о ведении их бизнеса, то сможете по этому анализу построить правильную стратегию временного таргетинга.
Этап 3. Проводим пост-анализ
Хороший маркетолог не думает — он тестирует!
Даже проведя весь необходимый первоначальный анализ, вы никогда не сможете выбрать время лучше, чем выберут его ваши покупатели. Именно поэтому очень важно, что бы у вас на сайте точно и без ошибок отслеживались макроконверсии. Не столь важно, как именно вы будете их отслеживать: через Analytics или Метрику, однако они обязательно должны учитываться.
Рис.3 В русскоязычной метрике введите в поиске «время дня» и найдите аналогичный отчет.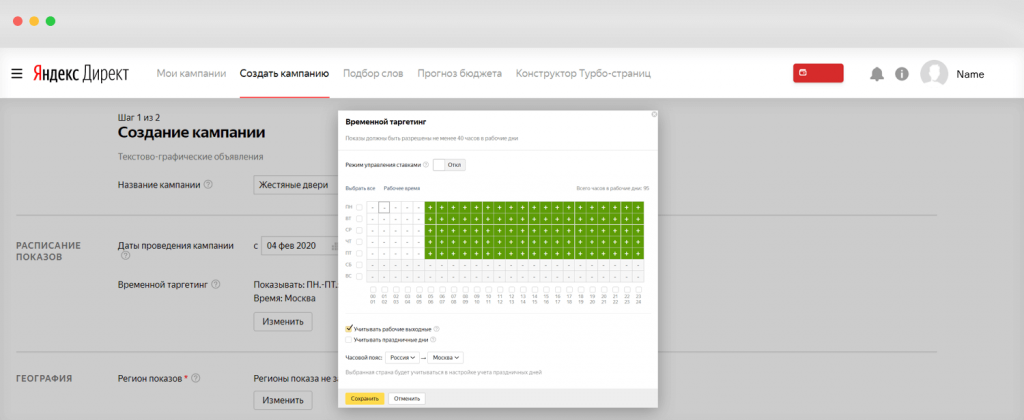
Спустя несколько недель после запуска вы сможете провести пост-анализ и выявить наиболее конверсионное время. Для этого я в данном примере буду использовать Яндекс.Метрику, т.к. она имеет большее распространение. В Яндекс.Метрике 2.0 необходимо зайти в отчет «трафик на сайт относительно времени» и выбрать сегмент пользователей, которые достигли ту или иную цель на вашем сайте. Пример указан на рис. 3.
Рис.4 На графике четко видны всплески активности регистраций в 12 часов дня, в 15, 17 и 21 час.
Помимо количества конверсий вы можете провести анализ конверсии в %, а также суммы полученных заказов, если у вас подключен модуль электронной торговли. В полученном отчете вы увидите зависимость ваших регистраций от времени суток. При этом обязательно учитывайте 2 важных фактора — зависимость регистраций от времени показа (не останавливается ли у вас днем реклама, за счет чего падает трафик), а также зависимость от среднего дня (в 3 часа ночи онлайн меньше, чем в 7 вечера). Вы сможете перераспределить ваши рекламные бюджеты исходя из наиболее конверсионных часов.
Вы сможете перераспределить ваши рекламные бюджеты исходя из наиболее конверсионных часов.
Подведем итог
В разное время одна и та же аудитория на одном и том же сайте может вести себя по-разному. Имея определенные ограничения по предельной стоимости заявки и суммарному дневному бюджету, наиболее правильным подходом будет перераспределение бюджета на наиболее эффективное время. Для выявления конверсионных часов существуют три основных метода:
- Анализ продукта и поведения аудитории
- Анализ конкурентов
- Пост-анализ конверсий на вашем сайте
Не забывайте, что количество конверсий может зависеть от разных факторов, в том числе от источника трафика и различных офферов. Не теструйте сразу несколько переменных!
Давайте дружить!
ОСТАВИТЬ ЗАЯВКУ!
Как таргетинг в режиме реального времени может революционизировать вашу маркетинговую стратегию: BDEX
Предоставление нужного контента в нужный момент является жизненно важной частью привлечения и удержания клиентов маркетинга и рекламы. Давно прошли времена распылительного маркетинга, когда рекламодатели рассылали и распространяли массовые кампании в надежде превратить несколько любопытных глазных яблок в платящих клиентов. Сегодняшние потребители узнают общую рекламу или сообщение, когда видят их, и полностью отключаются. Они требуют персонализации на протяжении всего клиентского опыта, и это начинается с актуальной и своевременной маркетинговой кампании.
Давно прошли времена распылительного маркетинга, когда рекламодатели рассылали и распространяли массовые кампании в надежде превратить несколько любопытных глазных яблок в платящих клиентов. Сегодняшние потребители узнают общую рекламу или сообщение, когда видят их, и полностью отключаются. Они требуют персонализации на протяжении всего клиентского опыта, и это начинается с актуальной и своевременной маркетинговой кампании.Благодаря таргетингу в режиме реального времени компании могут получать точные данные, которые им нужны, связываться со своими клиентами независимо от их устройства и проводить максимально эффективные маркетинговые кампании.
Поиск нужных данных в нужное время По своей сути маркетинг в реальном времени предоставляет нужный контент нужному человеку на нужном устройстве в нужное время. Конечно, это легче сказать, чем сделать, особенно если вы не знаете, когда и где ваши потенциальные клиенты делают покупки или какие продукты и услуги они хотят купить. Многие компании полагаются на обычные устаревшие ресурсы, которые предоставляют только традиционные данные или прогнозную аналитику. Хотя такая информация полезна для создания клиентской базы, она имеет ограниченную ценность, когда речь идет о маркетинге в реальном времени.
Многие компании полагаются на обычные устаревшие ресурсы, которые предоставляют только традиционные данные или прогнозную аналитику. Хотя такая информация полезна для создания клиентской базы, она имеет ограниченную ценность, когда речь идет о маркетинге в реальном времени.
Введите таргетинг в реальном времени. Помечая, собирая и интегрируя данные из онлайн- и офлайн-каналов, а также с различных мобильных и настольных устройств, компании могут получать информацию, необходимую им для таргетинга потенциальных клиентов в тот момент, когда они начинают делать покупки.
Сбор релевантных данных в режиме реального времениЧтобы компании могли быстро и точно найти потенциальных клиентов с помощью таргетинга в режиме реального времени, BDEX собирает данные в режиме реального времени: автомобили, одежда, мебель и др.).

Этот массив данных из разрозненных источников затем интегрируется в платформу обмена данными BDEX, где он становится уникальной, целенаправленной и полезной маркетинговой информацией, которую компании могут использовать для обеспечения и улучшения взаимодействия между людьми.
Подключайте клиентов с нескольких устройствОднако таргетинг в реальном времени — это не просто поиск нужных данных. Знание того, на кого и на какое устройство ориентироваться, является важным шагом в любой эффективной маркетинговой стратегии.
Распространение мобильных устройств позволило людям во всем мире получить доступ к Интернету, но также усложнило для маркетологов поиск клиентов на разных устройствах. Чтобы систематизировать огромное количество данных о своих клиентах, компании внедряют графы идентификаторов или базы данных, которые содержат профили клиентов и все известные идентификаторы, связанные с этими потребителями. Однако, если информация в этих базах данных основана исключительно на вероятности подключения пользователя к устройству, график идентификаторов может не обеспечивать максимально точное совпадение, а поскольку потребители постоянно меняются и включают новые устройства, это вызов, чтобы узнать, актуальны ли данные вашего устройства.
Чтобы обеспечить уверенность при сопоставлении клиентов с устройствами, граф идентификаторов BDEX использует детерминированный подход сопоставления между устройствами. Вместо того, чтобы полагаться на вероятностные показатели, BDEX использует детерминированную аутентифицированную информацию о клиентах, такую как анонимные данные для входа в систему, адреса электронной почты и покупки по кредитным картам, чтобы сопоставлять и распознавать людей независимо от того, какое устройство они используют. Благодаря информации, поступающей в режиме реального времени от более чем 75 проверенных партнеров и более чем 800 миллионам мобильных подключений к электронной почте, BDEX может помочь вам ориентироваться на потребителей на разных платформах с непревзойденной точностью.
Как только компании получат множество данных в режиме реального времени и смогут успешно сопоставлять пользователей с их правильными устройствами, они могут создавать маркетинговые кампании, адаптированные для конкретных клиентов и каналов.
Персонализация является ключевым компонентом эффективной маркетинговой кампании и успешных отношений с клиентами. Клиенты не только ценят персонализацию, но и ожидают ее. По данным Salesforce, 76% потребителей ожидают, что компании поймут их потребности и ожидания.
С помощью таргетинга в режиме реального времени и сопоставления разных устройств компании могут:
- Адаптировать маркетинговые сообщения на основе поведения. Например, если на прошлой неделе клиент искал пару обуви с помощью смартфона, компания может отправить персонализированный код купона, чтобы поощрить транзакцию.
- Предлагайте релевантные рекомендации по продуктам. Основываясь на недавних покупках клиента или недавней истории поиска, компания может предложить аналогичные продукты или другие предметы, которые дополнят покупку.
- Получите больше преимуществ от программной рекламы. Точные данные в режиме реального времени — основа успешной программной рекламы.
- Поощряйте покупателей приходить в магазин. Розничные продавцы, рестораны и другие предприятия могут использовать технологию геозоны, чтобы знать, когда их клиенты находятся рядом, и поощрять их посещение с помощью push-уведомлений, текстовых сообщений и других каналов.
Это всего лишь несколько способов использования данных в режиме реального времени для развития взаимодействия между людьми и повышения качества обслуживания клиентов. Используя персонализацию для привлечения клиентов и взаимодействия с ними, компании могут развивать подлинные отношения, которые приводят к увеличению продаж и укреплению лояльности к бренду.
Хотя эта тактика очень эффективна, 60% маркетологов пытаются персонализировать контент в режиме реального времени. Чтобы помочь компаниям в их маркетинговых усилиях и не только, BDEX разработала инструмент Explore Data Tool, который позволяет компаниям анализировать различные наборы данных, чтобы привлекать клиентов на протяжении всего процесса покупки: от создания индивидуальной аудитории до таргетинга в реальном времени и после покупки.
Благодаря более чем 100 партнерам аналитики BDEX могут искать более 1 триллиона точек данных о каждом потребителе в США в 19категории покупателей и более 5000 других категорий, призванных помочь маркетологам точно настроить маркетинг в реальном времени. Свяжитесь с нами по телефону , чтобы узнать, как мы можем помочь вам улучшить вашу текущую маркетинговую стратегию.
Правильные люди, нужное время, нужное место: ориентация на вашу аудиторию в Интернете
Преимущество онлайн-рекламы заключается в том, что бренды могут найти практически любого, с любым интересом и в любом сегменте, который они ищут. Например, если бизнес продает веганские кожаные кошельки и хочет ориентироваться на мужчин с располагаемым доходом, которые заинтересованы в экологически чистых продуктах, они могут их найти!
Удивительно, но благодаря прорывам в цифровом маркетинге у брендов появилась гибкость, чтобы охватить обширные группы клиентов, а затем, в свою очередь, нацелиться на определенные сегменты аудитории. Однако у такого уровня свободы есть следствие — дополнительный уровень сложности вашей маркетинговой стратегии. В «старые времена» маркетинга было достаточно создать броский, привлекательный креатив и разместить его в газете, на рекламном щите или на радио.
Времена изменились. С силой гипертаргетинга приходит ответственность использовать его с умом, вдумчиво определять аудиторию и общаться с ней персонализированным образом. Бренды, которые делают это хорошо, выделяются среди конкурентов, вызывают симпатию у своих клиентов и пользуются преимуществами выгодных долгосрочных отношений с клиентами.
Подумай об этом. Допустим, бренд продает высококачественные Bluetooth-динамики. Эти динамики серьезно технологически продвинуты и оснащены функциями, которые в полной мере оценят настоящие акустические системы. Они также поставляются с дизайном, который гармонирует с любой обстановкой от дома до офиса и многого другого. Они также оценены в соответствии с их высококлассным характером, начиная с 400 долларов.
Креатив кампании бренда (как статический, так и видео) украшен красивыми элегантными динамиками и имеет несколько преимуществ. Эти объявления связаны с целевой страницей, которая содержит всю необходимую информацию и упрощает покупку. Однако, поскольку у бренда мало информации о своей целевой аудитории, они настроили общий таргетинг без реальных ограничений по возрасту, полу, интересам, доходу и т. д.
Кампания продолжается какое-то время, и хотя результаты неплохие, бренд недоумевает, почему они не лучше. Они хотят увеличить рейтинг кликов (CTR), повысить посещаемость сайта, снизить показатель отказов и повысить рентабельность инвестиций, но не знают, как это сделать.
Решение заключается в тщательной разработке стратегии для конкретной целевой аудитории. Люди не хотят чувствовать себя продаваемыми — они хотят, чтобы с ними общались. Между этими двумя вещами есть большая разница, и ключ заключается в создании персонализированного опыта с помощью рекламы, целевых страниц и, да, определенного таргетинга.
Если что-то из этого найдет отклик, то эта статья будет вам по душе. В предыдущих разделах мы рассмотрим:
- Почему создание персонализированного опыта является ключом к лучшим результатам
- Как определить вашу целевую аудиторию
- Основные виды таргетинга и плюсы и минусы каждого
- Что делать дальше
Персонализированные рекламные услуги: что это такое и как их следует использовать?
Клиенты открыты для рекламы, пока реклама не является навязчивой, и они извлекают из нее некоторую ценность. Бюро интерактивной рекламы (IAB) обнаружило, что 3 из 4 пользователей хотят меньше, но более персонализированной рекламы. Бренды, которые инвестируют в персонализацию рекламы, в том числе в узконаправленную рекламу, имеют возможность создавать и развивать выгодные отношения со своими клиентами, которые со временем увеличивают пожизненную ценность (LTV).
Что означает персонализация?
Персонализация — это широкая тема, которая может включать электронную почту, сайты и рекламу. В данном случае мы имеем в виду создание в высшей степени персонализированной рекламы путем распределения целевых аудиторий по значимым сегментам и распространения рекламы в этих сегментах на основе их определяющих мотивов, характеристик и потребностей. Например, давайте посмотрим на бизнес, который продает чайники, чай и чайные принадлежности. Они могут разделить свою аудиторию на сегменты, которые представляют разные персонажи, например:
· Любитель чая на ходу: В основном этот человек пьет чай на работе или по дороге на работу и с работы. Этот человек пьет чай как более здоровую и низкокалорийную альтернативу газировке или латте. Скорее всего, это люди в возрасте от 25 до 50 лет, которые делают карьеру. Для этого человека вы бы нацелились на быстро и легко завариваемые сорта чая и дорожные кружки или варианты, удобные для рабочего стола. В своем креативе вы хотите подчеркнуть пользу для здоровья и удобство для путешествий.
· Медленный глоток: Этот человек наслаждается ежедневным ритуалом чаепития. Им нужен красивый чайник, красивая чашка и чашка хорошо заваренного чая. Этот сегмент на 75% состоит из женщин и на 25% из мужчин, как правило, имеет более высокий располагаемый доход, чем средний, и любит такие вещи, как йога, тай-чи, поэзия и походы. Вы нацеливаетесь на этого человека с помощью сообщений и продуктов, которые отражают его потребность в покое, размышлениях и чашке горячего чая.
Это лишь два из многих способов сегментации аудитории. Представьте себе сегменты аудитории специально для людей, заинтересованных в здоровье, просто женщин, интересующихся цветочными вариантами, людей, которые хотят зарядиться энергией от зеленого чая и т. д.
Эти две аудитории могут быть дополнительно сегментированы для создания более персонализированных впечатлений. Например, сегмент чаепитий на ходу может быть разделен на дорожные кружки и варианты, удобные для стола, дорожные кружки с вкладышами и варианты, характерные для чайных пакетиков, специальные варианты дизайна, как правило, более привлекательные для женщин, и те, которые более привлекательны для женщин. мужчины и т. д. Варианты поистине безграничны. В руках маркетолога определить, насколько детально он хочет ориентироваться, какие сегменты он хочет исследовать и какие продукты он хочет продвигать.
В данном случае мы имеем в виду создание в высшей степени персонализированной рекламы путем распределения целевых аудиторий по значимым сегментам и распространения рекламы в этих сегментах на основе их определяющих мотивов, характеристик и потребностей.
Как определить сегменты клиентов?
Теперь, когда ваш разум переполнен всеми возможностями для создания персонализированной рекламы, давайте рассмотрим несколько советов по созданию сегментов клиентов на основе данных.
В идеале, определить, кто будет клиентом, хотя бы предварительно, нужно на самых ранних стадиях развития бизнеса или продукта. Однако определение целевой аудитории — это не разовое мероприятие. Это то, что следует часто уточнять, используя данные, имеющиеся в распоряжении бренда. Вот некоторые способы определения аудитории:
- Google Analytics: Посмотрите, какие люди приходят на ваш сайт, какие страницы они посещают, какой контент находит отклик и т. д.
- Социальные сети: Следите за тем, кто взаимодействует с вашими публикациями, пишет о вас и что именно они говорят.
- Отзывы клиентов: Напрямую запрашивайте отзывы клиентов с помощью опросов и фокус-групп.
Если маркетинг — это построение отношений (подсказка: так и есть), то умение слушать — ключ к успеху. Создание систем, сосредоточенных на том, чтобы прислушиваться к клиентам, узнавать о них больше и применять то, что вы узнали, почти ничего не будет стоить, но принесет большие дивиденды в виде вашей способности строить прибыльные долгосрочные отношения с клиентами.
Таргетинг 101
Итак, вы определили несколько идей для аудитории и подготовили персонализированный креатив. Теперь пришло время настроить высокотаргетированные кампании.
Сначала кратко о том, как работает таргетинг. Будь то размещение рекламы через Facebook, Instagram или Google, эти платформы могут определить довольно конкретное представление о том, кто использует устройство. Они могут определить возраст, пол, местоположение и даже интересы человека, отслеживая страницы, которые он посещает, вещи, которые он покупает, его публикации в социальных сетях, лайки и многое другое. Ничто из этого не является полностью ошибочным, но алгоритмы, которые создают профили для людей, довольно сложны.
Эти профили разбиты на множество атрибутов. Итак, в общих чертах профиль может выглядеть примерно так:
- Женщина
- 25-35
- Живет в районе метро Чикаго .
- Любит бегать
- Имеет маленьких детей
- Часто ищет пиццерии
- Ранее купленная футболка фаната Гарри Поттера
Каждый профиль представляет собой смесь демографических данных (пол, возраст, местонахождение) и интересов (хобби, семья, еда).
Примечание. это просто очень простое объяснение, которое подходит для наших целей в этой статье.
Помимо демографического таргетинга и таргетинга по интересам, маркетологи также могут нацеливаться на контекст и на похожую аудиторию. Ниже приводится объяснение каждого типа таргетинга, где он полезен, а также его уникальные недостатки.
Демографический
Это одна из самых основных форм таргетинга, которая включает в себя параметры для географического региона, возраста, пола, годового дохода и т. д. Ориентация на аудиторию с использованием демографических данных довольно проста. Как правило, бренд создает продукты, ориентируясь на определенного клиента, а это означает, что они снабжены информацией для создания одного или двух основных демографических профилей для аудитории на рынке.
Основным недостатком демографического таргетинга является то, что существует огромное разнообразие даже среди людей, которые выглядят одинаково демографически. Это затрудняет создание по-настоящему персонализированного опыта, потому что аудитория все еще может быть довольно широкой.
Интерес
Таргетинг по интересам все еще может быть простым (просто выберите некоторые интересы, которые есть у целевой аудитории), но требует более предусмотрительного и стратегического планирования. Хотя вы, вероятно, можете определить некоторые основные интересы целевой аудитории прямо за воротами, чуть более глубокое изучение данных может привести к некоторым интересным выводам.
Недостаток таргетинга по интересам очень важен — проблемы конфиденциальности. Мы все слышали новости и видели, как вступили в силу такие законы, как Общий регламент по защите данных (GDPR) и Калифорнийский закон о конфиденциальности потребителей (CCPA). Хотя будущее этих вариантов таргетинга неясно, кажется очевидным, что по крайней мере некоторая аудитория будет недоступна либо по демографическим параметрам, либо по интересам.
Контекстный
Контекстный таргетинг — идеальный вариант для брендов, которые хотят контролировать, где размещается их реклама в Интернете. Бренды могут обращаться к клиентам на сайтах, которые уже посещает их целевая аудитория, и на сайтах, которые совпадают с их ценностями. Кроме того, реклама может быть исключена с определенных веб-сайтов, которые считаются неприемлемыми. Это идеальный вариант для бизнеса, заботящегося о конфиденциальности, который подпадает под ограничения CCPA или GDPR.
Недостаток контекстного таргетинга в том, что он немного менее конкретен, чем другие, и может также потребовать дополнительных исследований, чтобы точно определить, какой тип контента или сайтов находит отклик у аудитории.
Двойник
Другие варианты предполагают проведение исследований, выбор альтернатив, их мониторинг и соответствующую оптимизацию. Lookalike таргетинг не требует ничего из этого. Бренды могут рассчитывать на то, что технология ИИ сделает за них тяжелую работу.
Каждая платформа по-своему подходит к таргетингу на двойников, но основное объяснение заключается в том, что система анализирует текущий трафик веб-сайта, тех, кто заходит на сайт, и тех, кто с наибольшей вероятностью совершает конверсию или выполняет какую-либо конкретную цель. Затем ИИ выходит и идентифицирует людей, похожих на текущих клиентов, и показывает им рекламу. С этой опцией он учится по ходу дела и приспосабливается к любым изменениям в продукте, трафике и т. д.
Единственным недостатком двойного таргетинга является то, что он не дает большого контроля над тем, на кого он направлен. Это, в свою очередь, не позволяет вам использовать таргетинг для тестирования новых аудиторий, которые могут выглядеть или вести себя иначе, чем те, на которые вы сейчас ориентируетесь.
Двойник не требует ничего из этого. Бренды могут рассчитывать на то, что технология ИИ сделает за них тяжелую работу.
Обобщенный таргетинг
Каждая из этих форм таргетинга имеет сильные и слабые стороны, которые следует учитывать при построении любой рекламной кампании. Таким образом, в большинстве случаев брендам рекомендуется использовать комбинацию всех этих вариантов. Используйте демографический таргетинг, таргетинг по интересам и контексту, чтобы выбрать конкретную аудиторию, и позвольте сложной технологии искусственного интеллекта, лежащей в основе двойного таргетинга, определить аудиторию, которая в противном случае могла бы быть упущена.
Что дальше?
Теперь, когда вы понимаете основы идентификации, поиска и общения с идеальной аудиторией, пришло время воплотить все эти идеи в жизнь. Ниже приведены некоторые полезные ресурсы, которые помогут определить аудиторию, создать активы кампании, которые будут резонировать с ними, измерить ваше влияние и оптимизировать ваши кампании.
Кто ваш клиент?
- Руководство по созданию надежного профиля идеального клиента
- В поисках мистера(ов) справа: 5 способов, как профиль идеального клиента улучшает праздничные продажи
- 5 инструментов, которые помогут вам узнать о вашей целевой аудитории
- Как создать профиль аудитории
- 7 способов получить преимущество благодаря анализу конкурентов
Вдохновение для кампании
- Не упустите свой шанс: произведите хорошее впечатление с помощью видеоконтента
- Как измерить успех видеоконтента
- Причины, по которым вы не используете видео (и почему вы должны)
- 4 компании D2C правильно рассказывают истории о брендах
Измерение и оптимизация
- Как разработать мультисенсорную модель атрибуции
- Взлом кода целей кампании и эффективности рекламы с атрибуцией
Первоначально опубликовано 2 октября 2019 г. , последнее обновление — 16 сентября 2022 г.
Таргетинг на аудиторию — охватите своих клиентов
Ваша аудитория слушает.
Когда люди погружаются в музыку и подкасты на Spotify, они доверяют нам, что знают их вкус и правильно используют их алгоритм. То же самое относится и к рекламе, которую они видят на нашей платформе. Наши инструменты таргетинга на аудиторию доносят ваше сообщение до людей в моменты, актуальные для них и для вашего бизнеса.
Узнайте о наших возможностях
С помощью целевой онлайн-рекламы вы получите доступ к инструментам поиска, которые свяжут вас с вашей аудиторией, где бы она ни находилась. Возьмите под контроль свои следующие маркетинговые кампании, поднимите свою компанию и добейтесь большей рентабельности инвестиций на основе заданных параметров.
Откройте для себя возможности маркетинга аудитории и улучшите свою цифровую стратегию таргетинга.
Начнем с основ. Свяжитесь со своей целевой аудиторией, используя собственные демографические данные Spotify.
- Возраст
- Пол
- Местонахождение
- Языки
Вы можете многое узнать о человеке по тому, что он слушает. Выйдите за рамки демографических данных, охватывая свою целевую аудиторию с помощью поведения при прослушивании, которое дает уникальное представление о том, чем они занимаются.
- Интересы
- База поклонников
Определите среду и контекст, в которых показываются ваши рекламные объявления. Быть услышанным (и увиденным) в моменты, связанные с опытом слушателя.
- Платформы и устройства
- Музыка
- Жанры
- Контексты в реальном времени
- Подкасты
- Темы выпусков подкастов
О, ты фантазируешь, да? В настоящее время мы предлагаем варианты пользовательского таргетинга только для рекламодателей, работающих с нашим отделом продаж с полным спектром услуг. Свяжитесь, чтобы узнать больше.
Реклама на Spotify — это не просто реклама.
Они часть шоу.Как рекламодатель вы участвуете в прослушивании музыки Spotify. Плавно вплетайте свою рекламу в любой плейлист или подкаст и продавайте свои продукты слушателям, не портя их погружения. Таргетинг на аудиторию никогда не был таким простым благодаря нашим возможностям целевого маркетинга.
Настройте все, от времени суток до почтового индекса, в котором живет ваша целевая аудитория. Реклама Spotify — это не просто реклама; они нацелены на толпу.
Создайте рекламу, соответствующую моменту.
Наши творческие инструменты самообслуживания упрощают создание аудиорекламы, соответствующей вкусам и контексту прослушивания вашей целевой аудитории, в нужный момент. Целенаправленная цифровая реклама гарантирует, что ваши объявления будут восприниматься как часть шоу, а не как вторжение.
Прочтите наше руководство по аудиотворчествуБудьте увиденными и услышанными.
Используйте форматы аудио- и видеорекламы, чтобы заинтересовать целевую аудиторию, когда они используют приложение Spotify или проводят время без экрана. Смешивайте и подбирайте свои форматы, чтобы ваш бренд чувствовал себя свежим каждый раз, когда его видят или слышат. Сделайте опыт специфичным для этого продукта и этого формата.
Тестируйте, учитесь, оптимизируйте, повторяйте.
Не знаете, какие инструменты таргетинга использовать? Протестируйте различные сегменты и подходы к таргетингу, чтобы увидеть, что лучше всего подходит для вашего сообщения. От таргетинга на аудиторию до таргетинга на СМИ — существует множество способов раскрыть свой потенциал как бренда. Мы помогаем вам получить максимальную отдачу от вашего рекламного бюджета и оставаться впереди конкурентов.
Прочтите наше руководство по оптимизации кампанииУзнайте, как эти рекламодатели использовали наши инструменты таргетинга для масштабирования охвата и максимизации результатов кампании.
Часто задаваемые вопросы по таргетингу на аудиторию
Платформа Spotify для таргетинга на аудиторию проста в освоении и использовании, но всегда есть вопросы, на которые каждый маркетолог хочет знать ответ, прежде чем приступить к работе. Вот некоторые из наиболее часто задаваемых вопросов.
Кампания, в основе которой лежит таргетинг на аудиторию, даст вам больше шансов эффективно привлечь людей, которые уже заинтересованы в ваших услугах. Целевой онлайн-маркетинг гарантирует, что вашу рекламу увидят нужные люди — независимо от того, ориентируетесь ли вы на представителей аудитории на основе их возраста, пола, местоположения или интересов, которые делают их совместимыми с вашим брендом.
Никто лучше вас не знает вашу аудиторию. Вы знаете, как они ведут себя на вашем веб-сайте, как взаимодействуют с вами в социальных сетях и как ведут себя офлайн. Это бесценное знание, которое должно стать основой вашей кампании по таргетингу, и чтобы получить наилучшие результаты от высокоточных инструментов таргетинга Spotify, вам нужно тестировать, тестировать и тестировать снова.
Частое тестирование поможет вам точно определить свою аудиторию и донести сообщение, которое найдет у них наибольший отклик.
Короче говоря, нет. Платформа самообслуживания Spotify предназначена для всех, кто управляет или представляет бизнес и нуждается в эффективных рекламных решениях, независимо от того, является ли он экспертом в области маркетинга или нет. Наши инструменты просты в использовании, и наша команда всегда готова ответить на любые ваши вопросы о проведении ваших кампаний.
Опередите конкурентов и заговорите с потенциальными клиентами, приступив к разработке своей первой рекламной кампании.
Готовы достучаться до своей аудитории в их счастливом месте?
Зарегистрируйтесь или войдите в свою учетную запись Ad Studio, чтобы создать кампанию. Или свяжитесь с нами напрямую, используя форму ниже.
Обзоры целевых показателей времени: цены и функции программного обеспечения — 2022 г.
#16 Индекс популярности
ОЦЕНКА HT
9 из 100
Hotel Tech Score — это составной рейтинг, состоящий из ключевых сигналов, таких как: удовлетворенность пользователей, количество отзывов, давность отзывов и информация, предоставленная поставщиком, чтобы помочь покупателям лучше понять их продукты.
Узнать больше
Сводка отзывов
16-е место из 25 в категории «Планирование и управление трудовыми ресурсами»
Простота использования
3,0/5
рентабельность инвестиций
4,0/5
Реализация
4,0/5
Служба поддержки
3,0/5
Процессы поддержки клиентов TimeTarget еще не проверены Hotel Tech Report.
Узнать больше
Самый популярный для
TimeTarget — австралийская компания-разработчик программного обеспечения, которая гордится тем, что предлагает рынку ведущее программное решение для учета рабочего времени и посещаемости/управления рабочей силой. Наша команда разработчиков из Сиднея следит за тем, чтобы TimeTarget постоянно улучшался за счет обновлений и дополнительных пользовательских функций, разработанных в ответ на потребности клиентов. Всегда прислушивались к нашим клиентам и старались…
Основана в 2002 году | Штаб-квартира в Милсонс-Пойнт, Австралия | 91 сотрудник
1/1
TimeTarget не имеет проверенных интеграций. Загляните позже, чтобы узнать о новых интеграциях TimeTarget.
Читать отзывы
Посмотреть альтернативы
Посмотреть альтернативы
УниФокус
Планирование и управление трудовыми ресурсами
Интегрированные инновационные инструменты управления персоналом, облегчающие управление трудовыми ресурсами, учет рабочего времени и посещаемости, соблюдение нормативных требований и многое другое
Посмотреть альтернативы
Посмотреть альтернативы TimeTarget
Найти идеальный вариант
Хотите узнать, подходит ли TimeTarget лучше всего для вашего отеля?
пройти викторину
НАЧАЛО РАБОТЫ
Загрузить руководство покупателя по планированию и управлению трудовыми ресурсами на 2021 год
Бесплатная загрузка
-
Переключение смен
ВАЖНАЯ ФУНКЦИЯЕсли вы потеряете члена команды, найдите квалифицированную замену одним нажатием кнопки.
Или сэкономьте свое время и позвольте сотрудникам поменяться сменами между собой. -
Автоматическое планирование
ВАЖНАЯ ФУНКЦИЯИспользуйте искусственный интеллект, чтобы предсказать, сколько сотрудников вам потребуется для удовлетворения ожидаемого спроса, и создавайте графики работы, чтобы соответствовать в кратчайшие сроки.
-
Приложение часов
ВАЖНАЯ ФУНКЦИЯТочно записывайте, когда и где работают ваши люди, чтобы получать точные табели учета рабочего времени, обновления посещаемости в режиме реального времени и быстрый и точный расчет заработной платы.
-
Динамическое планирование
ВАЖНАЯ ФУНКЦИЯ -
Трудовые нормы
ВАЖНАЯ ФУНКЦИЯ
-
Сравнительный анализ
ВАЖНАЯ ФУНКЦИЯ -
Отчетность и аналитика
ВАЖНАЯ ФУНКЦИЯ -
Корпоративная видимость
ВАЖНАЯ ФУНКЦИЯ -
Автоматизированный мониторинг труда
ВАЖНАЯ ФУНКЦИЯ -
Трудовые нормы
ВАЖНАЯ ФУНКЦИЯ -
Интеграция прогноза занятости
ИННОВАЦИОННАЯ ОСОБЕННОСТЬ
-
Командная связь
ВАЖНАЯ ФУНКЦИЯДелитесь важной информацией и документами с одним сотрудником или целыми командами и получайте уведомления, когда важные сообщения прочитаны.
-
Мобильное приложение
ВАЖНАЯ ФУНКЦИЯУправляйте своими командами с любого устройства и упростите для сотрудников просмотр еженедельных расписаний, обновлений и задач.
ОБСЛУЖИВАЕМЫЕ РЫНКИ
- Северная Америка
- Южная Америка
- Европа
- Центральная Америка
- Азиатско-Тихоокеанский регион
- Средний Восток
- Африка
- Антарктида
- Океания
ПРОВЕРЕННЫЕ РЫНКИ
Вероятность рекомендовать
Фильтровать по типу недвижимости
Фильтровать по географии
Оповещение о низком объеме отзывов
Этот продавец на 100% ниже среднего по категории.
Снижение доли голоса
У этого поставщика меньше недавних отзывов, чем в среднем, по сравнению с другими ведущими поставщиками планирования и управления трудовыми ресурсами. Перед продолжением рекомендуется запросить последние рекомендации.
Что тебе больше всего нравится?
Хороший реестр и система обмена сообщениями для персонала
Что вам не нравится или что можно было бы улучшить?
Обслуживание клиентов плохое, и система может работать очень медленно
Посмотреть все
Планирование и управление трудовыми ресурсами
Индекс затрат на оплату труда в отелях и отчет об анализе уборки
Опубликовано Эффективность отеля
Планирование и управление трудовыми ресурсами
Инструментарий совершенствования труда: 4-этапный план достижения успеха в трудовых отношениях
Опубликовано Эффективность отеля
Посмотреть все
Популярные интеграции целевого времени
О-о… похоже, у Time Target нет в списке интеграций.
Повернуть время вспять: воздействовать на инфильтрирующие опухоль миелоидные клетки, чтобы остановить прогрессирование рака
Введение в роль миелоидных клеток в развитии рака
До начала двадцать первого века рак считался заболеванием, поражающим одну клетку. Этот упрощенный взгляд уступил место новому взгляду. По мере развития опухоли происходят неустанные перекрестные взаимодействия между гетерогенными злокачественными клетками и соседними паренхиматозными, стромальными и рекрутированными иммунными клетками. Эти клетки вместе с внеклеточным матриксом и растворимыми медиаторами образуют микроокружение опухоли (МОО). Состав иммунного инфильтрата в ТМО во многом определяет прогрессирование рака и чувствительность рака к различным видам терапии. Опухолепроникающий CD8 + Т-лимфоциты играют ключевую роль в борьбе с раковыми клетками. Следовательно, активация de novo или существующих CD8 + Т-клеток с использованием таких стратегий, как противораковые вакцины и терапия иммунных контрольных точек, привлекла значительное внимание (1). Устойчивые ответы, полученные с помощью этих методов лечения, побудили использовать иммунотерапию рака в стандарте лечения различных типов рака. Несмотря на эти огромные улучшения в лечении рака, значительное число пациентов не получают пользы от современной иммунотерапии рака. Существует достаточно доказательств того, что миелоидные клетки, присутствующие в TME, лежат в основе неудачи этой терапии. Таким образом, инфильтрирующие опухоль миелоидные клетки (TIM) считаются важными терапевтическими мишенями (2).
Миелоидные клетки представляют собой гетерогенную группу иммунных клеток, принадлежащих врожденной иммунной системе. Среди миелоидных клеток большое внимание уделяется моноцитам, макрофагам, дендритным клеткам (ДК) и гранулоцитам. Эти клетки, каждая по-своему, играют существенную роль в тканевом гомеостазе. Более того, моноциты, макрофаги и ДК хорошо известны своей способностью регулировать ответы Т-клеток, тем самым объединяя врожденный и адаптивный иммунитет. Опухолевые клетки используют преимущества миелоидных клеток для поддержания тканевого гомеостаза, используя способность миелоидных клеток продуцировать медиаторы воспаления [например, интерлейкин-6 [ИЛ-6] и фактор некроза опухоли-α [ФНО-α]], факторы роста, которые влияют на пролиферация опухоли и образование сосудов [например, трансформирующий фактор роста-β [TGF-β] и фактор роста эндотелия сосудов [VEGF]], а также ферменты, расщепляющие матриксные белки [например, матриксные металлопротеиназы [MMPs]] (3, 4) . Более того, опухолевые клетки используют способность миелоидных клеток контролировать ответы Т-клеток. Таким образом, инструктируемые опухолевыми клетками, миелоидные клетки помогают в создании TME, который характеризуется хроническим воспалением, иммуносупрессией и непрерывно пролиферирующими опухолевыми клетками, которые могут диссеминировать. Это важные признаки рака (5). Как это ни парадоксально, миелоидные клетки также участвуют в разрешении рака (4). Миелоидные клетки могут выполнять глубокие противоопухолевые функции, такие как прямое уничтожение опухолевых клеток наряду с непрямым уничтожением опухолевых клеток посредством активации, среди прочего, CD8 + Т-клетки. В оставшейся части введения мы обсуждаем про- и противоопухолевые свойства различных подмножеств TIM в контексте парадигмы иммуноредактирования рака (6). Кроме того, мы обсуждаем TIM и то, как они влияют на различные методы лечения рака. Наконец, мы представляем обзор подходов, которые были изучены для нацеливания на TIM и, как таковые, повышения эффективности современных методов лечения рака.
Развитие и фенотип инфильтрирующих опухоль миелоидных клеток в двух словах
Опухолеинфильтрирующие миелоидные клетки (ТИМ) представляют собой гетерогенную популяцию клеток, характеризующихся разнообразием и пластичностью. Многие ТИМ происходят из циркулирующих моноцитов и гранулоцитов, которые, в свою очередь, происходят из гемопоэтических стволовых клеток костного мозга (ГСК; рис. 1). В отсутствие сигналов активации постоянная стимуляция опухолевыми факторами побуждает предшественников моноцитов и гранулоцитов отклоняться от их внутреннего пути терминальной дифференцировки в зрелые макрофаги, ДК или гранулоциты. Вместо этого предпочтение отдается дифференцировке в патологические, альтернативно активированные незрелые миелоидные клетки. Эти незрелые миелоидные клетки включают опухолеассоциированные DCs (TADCs), опухолеассоциированные нейтрофилы (TANs), клетки-супрессоры миелоидного происхождения (MDSCs) и опухолеассоциированные макрофаги (TAMs). В качестве альтернативы этому «аварийному миелопоэзу» ТАМ могут происходить из резидентных в тканях макрофагов, которые, в свою очередь, могут иметь эмбриональное или моноцитарное происхождение (7–9). ). Эти резидентные в тканях макрофаги претерпевают изменения фенотипа и функции во время канцерогенеза, и пролиферация, по-видимому, является ключом к поддержанию ТАМ, полученных из резидентных в тканях макрофагов.
Рисунок 1 . Прогрессирование от HSC к TIM, способствующему развитию опухоли. Отдельные этапы перехода от HSC к TIM происходят в разных местах и начинаются с амплификации и дифференцировки HSC и их предшественников, включая общий миелоидный предшественник (CMP), гранулоцитарно-моноцитарный предшественник (GMP), миелобласт (MB) и моноцитарно-дендритных клеток-предшественников (MDP) в костном мозге. Новые миелоидные клетки высвобождаются в кровоток, готовые мигрировать в ложе опухоли. Этот процесс регулируется молекулярными сигналами, продуцируемыми раковыми клетками, и дополнительно усиливается молекулярными сигналами, продуцируемыми, среди прочего, ТИМ. Эти факторы включают гранулоцитарный (G) и гранулоцитарно-макрофагальный (GM) колониестимулирующий фактор (CSF), Fms-подобную тирозинкиназу 3-лиганд (Flt3-L), хемокиновый (мотив CC) лиганд 2 (CCL2), VEGF и S100A8/9. . Показаны фенотип ТИМов человеческого и мышиного происхождения и их функциональные признаки. Для TADC, TAN и TAM рисунок упрощается, поскольку для этих типов клеток различают разные подмножества с анти- (стимулирующими) или проопухолевыми (регуляторными) функциями. На рисунке основное внимание уделяется подмножествам с проопухолевой активностью.
Среди TIM большое внимание привлекли MDSC. Эта популяция TIM была разделена на полиморфноядерные (PMN) и моноцитарные (M) MDSC, которые морфологически и фенотипически напоминают TAN и моноциты соответственно (10, 11). Наиболее важным критерием, определяющим MDSC, является их способность подавлять Т-клетки. Поскольку опухоль является почвой для опухолевых миелоидных клеток, термин MDSCs и его узкое разделение на PMN- и M-MDSCs является спорным (3). Два наблюдения говорят в пользу более широкого термина, объединяющего незрелые миелоидные клетки под одним зонтиком. Во-первых, было описано несколько условий, способствующих быстрой дифференцировке M-MDSC в PMN-MDSC, TAM или TADC, что подчеркивает пластичность миелоидных клеток и их способность изменять форму в зависимости от встречающихся сигналов (12–14). Во-вторых, характерная черта, подавляющая Т-клетки, не уникальна для MDSC. Также моноциты, дендритные клетки и макрофаги могут приобретать Т-клеточно-супрессивный фенотип при ТМЭ (3, 15). Приведенные выше результаты затрудняют разделение пластикового компартмента ТИМ на дискретные подмножества, но заставляют нас признать тот факт, что опухоль управляет своим окружением в соответствии со своими собственными правилами и формирует ТИМ, которые функционально участвуют в перекрестных диалогах, чтобы служить опухоли. Тем не менее, некоторые подмножества TIM могут проявлять функции, которые препятствуют прогрессированию опухоли, например, активация CD8, отторгающих опухоль.0665 + Т-клетки. Поэтому ранее мы предложили разделить TIM в соответствии с их функцией на миелоидные стимулирующие клетки, связанные с иммунной активацией, и миелоидные регуляторные клетки, связанные с иммуносупрессией и заживлением ран (3).
Функциональная поляризация TIM в сторону помощи или противодействия росту опухоли часто объясняется с помощью Т-хелперов (T H ) 1/2 парадигмы CD4 + Т-клеток. В частности, ТАМ и ТАН были подразделены на так называемые ТИМ 1-го и 2-го типов, которые, как таковые, классифицируют их как ТИМ, которые проявляют противоопухолевую или проопухолевую активность соответственно (16–21). Хотя деление на тип 1 и тип 2, безусловно, представляет интерес, следует подчеркнуть, что любая форма строгой классификации не отражает сложности in vivo ситуация. In vivo TIM подвергаются воздействию множества экологических стимулов, которые могут давать противоположные сигналы, что приводит к смешанным профилям, а не к строгому профилю типа 1 или типа 2. Это хорошо задокументировано для ТАМ (22–25).
Зрелые TADC и ТАМ 1 типа (TAM1) считаются иммуностимулирующими ТИМ. Оба характеризуются высокой экспрессией антигенпрезентирующих молекул (например, MHC-I и -II), костимулирующих молекул (например, CD80 и CD86), наряду с секрецией высоких уровней стимулирующих цитокинов (например, IL-6, IL -12, IL-23 и TNF-α) (24). Напротив, MDSC, TAM 2 типа (TAM2) и толерогенные TADC (tolDC) считаются иммунорегуляторными TIM. Они имеют много общих функциональных особенностей, которые обеспечивают сильную иммуносупрессию, среди которых экспрессия различных ферментов, таких как индуцибельная синтазы оксида азота (iNOS), аргиназа-1 (ARG-1) и индоламин-2,3-деоксигеназа (IDO), секреция цитокинов, таких как IL. -10 и TGF-β, а также экспрессию совместно ингибирующих молекул, таких как лиганд запрограммированной смерти 1 (PD-L1; рисунок 1) (3). Помимо иммуносупрессии, эти иммунорегуляторные TIM ответственны за ремоделирование тканей, стимулирование пролиферации опухолевых клеток и ангиогенеза, тем самым способствуя прогрессированию рака различными путями.
Проопухолевые признаки TIM используются большинством опухолей посредством культивирования регуляторных TIM до тех пор, пока, среди прочего, не будут установлены большие коллекции TAM2, MDSC, TAN2 и небольшие количества незрелых tolDC. Это устанавливает петли прямой связи, поскольку многие факторы, создаваемые регуляторными ИМП, обеспечивают создание, набор и функционирование новых регуляторных ИМП. Эти факторы включают VEGF, TGF-β, белки S100A8/9 и так далее. Поскольку проопухолевые пути и механизмы, используемые регуляторными TIM, демонстрируют избыточность, воздействия только на одно подмножество или путь, вероятно, будет недостаточно, чтобы остановить прогрессирование рака. В следующих разделах мы более подробно обсудим деятельность ТИМов в рамках парадигмы иммуноредактирования (6).
Миелоидные клетки имеют связь кошек и мышей с раковыми клетками
Иммуноредактирование является результатом динамического процесса сложных взаимодействий между иммунными и раковыми клетками. В этом процессе зарождающиеся трансформированные клетки первоначально распознаются и элиминируются различными клетками врожденной и адаптивной иммунной системы. Эта непрерывная иммунная атака приводит к отбору трансформированных клеток с неиммуногенными признаками, которые могут уклоняться от иммунной атаки. Это предвещает фазу равновесия, в которой злокачественные клетки сосуществуют с иммунными клетками. Некоторые из них проявляют противоопухолевую активность, в то время как другие проявляют активность, способствующую развитию опухолей. Наконец, раковые клетки больше не распознаются и не убиваются иммунной системой, и, таким образом, ускользают, чтобы прогрессировать в полномасштабную злокачественную опухоль. Миелоидные клетки участвуют в каждой фазе этого процесса (рис. 2, 3).
Рисунок 2 . Миелоидные клетки и их роль в элиминации раковых клеток. Миелоидные клетки в дополнение к другим типам иммунных клеток выполняют уникальные, а также избыточные функции для достижения элиминации опухолевых клеток. Зарождающиеся раковые клетки обнаруживаются NK-клетками посредством экспрессии специфических лигандов, например, лигандов для рецептора NKG2D. Это приводит к опосредованному NK-клетками уничтожению раковых клеток, которое может быть дополнительно усилено за счет активации макрофагов и DC за счет связывания их рецептора dectin-1 со структурами N-гликана, экспрессируемыми на определенных опухолевых клетках. Фрагменты раковых клеток, а также раковые клетки, экспрессирующие поверхностный кальретикулин, могут поглощаться макрофагами (при условии, что SIRPα не активирован) и ДК. Таким образом, эти антигенпрезентирующие клетки приобретают ТАА и могут активировать CD4 + и CD8 + Т-клетки. IFN-γ, продуцируемый этими Т-клетками, а также NK-клетками, является одним из механизмов, используемых для уничтожения раковых клеток. Это уничтожение раковых клеток может быть дополнительно усилено активацией онкоцидной программы (высвобождение NO и TNF-α) макрофагов через IFN-γ.
Рисунок 3 . Основные роли, которые играют миелоидные клетки при спасении раковых клеток от иммуноопосредованного разрушения. Растворимые факторы, секретируемые опухолевыми и иммунными клетками, такие как CCL2, IL-4, IL-6, IL-8, IL-10, IL-13, GM-CSF, M-CSF, VEGF и SCF, создают TME, в который прибывают и локальные миелоидные клетки превращаются в иммуносупрессивные ТИМ. Из них наиболее распространены TAM2, TAN2 и MDSC, в то время как tolDC менее распространены в TME. Цитокины, такие как IL-10 и TGF-β, в значительной степени способствуют созданию иммуносупрессивного TME. Кроме того, эти TIM экспрессируют множество ферментов (например, IDO, ARG-1, iNOS) и поверхностных маркеров (например, CD40, CD80, PD-L1), которые помогают им подавлять противоопухолевый иммунитет. Более того, ТИМ не только инструктированы подавлять противоопухолевые ответы под влиянием факторов роста и цитокинов в ТМО, но и выполнять действия, соответствующие потребностям опухолевых клеток. Эти действия включают поддержание хронического воспаления, стимулирование неоангиогенеза, рост опухоли, инвазию и метастазирование. Эти активности опосредованы секрецией растворимых факторов (например, цитокинов, факторов роста и протеаз). Они не показаны на рисунке.
Фаза элиминации
Наши знания о событиях, происходящих на стадии элиминации процесса иммуноредактирования, в значительной степени получены из исследований на модели рака на мышах. Они показывают, что интактный компартмент лимфоидных иммунных клеток играет центральную роль в принципе элиминации раковых клеток. Роль миелоидных клеток в элиминации зарождающихся трансформированных клеток изучается реже, и количество публикаций по этому вопросу ограничено. Тем не менее, несколько передовых исследований показывают, что миелоидные клетки вносят значительный вклад в эту стадию процесса иммуноредактирования.
Трансформированные клетки могут экспрессировать лиганды для рецептора естественного киллера (NK) группы 2 члена D (NKG2D), который у мышей экспрессируется на NK-клетках, CD8 + T-клетках и активированных макрофагах (26, 27), в то время как у людей его экспрессия ограничена NK-клетками и CD8 + Т-клетками (28). Связывание NKG2D с NK-клетками устанавливает его цитолитические функции (29). Другими рецепторами, экспрессируемыми на NK-клетках, которые участвуют в распознавании и уничтожении опухолевых клеток, являются NKp30, NKp44 и NKp46 (30) и CD226 (DNAM-1) (31). Также потеря MHC-I, которая часто происходит в раковых клетках (32), запускает цитолитические свойства NK-клеток, таких как рецепторы, такие как Ly49. который подает ингибирующие сигналы NK-клеткам, больше не задействованы. Исследования показывают, что NK-клетки со сниженной экспрессией Ly49 неспособны отторгать трансплантированные опухоли, предполагая, что правильное функционирование NK-клеток является ключом к отторжению опухоли in vivo (33). Макрофаги и ДК могут усиливать опосредованную NK-клетками цитолитическую активность в отношении опухолевых клеток (34, 35). В ходе экспериментов по совместному культивированию было установлено, что некоторые опухолевые клетки экспрессируют структуры N-гликанов, которые могут связывать dectin-1 на макрофагах и DC, что приводит к последующей активации регуляторного фактора интерферона 5 (IRF5) (35). Активность IRF5 была критической для индукции экспрессии INAM, мембраносвязанного белка, который, как известно, активирует NK-клетки посредством гомофильного взаимодействия (36).
Кроме того, хорошо известно, что трансформированные клетки экспрессируют антигены, образующиеся в результате мутаций, эпигенетических изменений или аберрантного процессинга белков. Эти ассоциированные с опухолью антигены (TAA) представлены на молекулах MHC-I и могут распознаваться Т-клетками CD8 + (37). Эти CD8 + Т-клетки могут получать костимуляцию посредством запуска их рецептора NKG2D лигандами, экспрессируемыми на трансформированных клетках (38). Далее было высказано предположение, что отторгающие опухоль CD8 + Т-клетки стимулируются ДК и макрофагами, которые поглощают фрагменты раковых клеток-мишеней NK-клеток (39). Более того, раковые клетки могут экспрессировать поверхностный кальретикулин в ответ на стресс (40). Поверхностная экспрессия кальретикулина служит сигналом «съешь меня», позволяя DC приобретать TAA, которые могут быть перекрестно представлены CD8 + Т-клеткам. Было показано, что для инициации такого противоопухолевого ответа CD8 + Т-клеток требуется эндогенный IFN I типа, поскольку мыши, у которых CD8α + ДК без IFNAR не способны индуцировать опосредованное лимфоцитами отторжение опухоли (41). Этот эндогенный IFN типа I может быть результатом реакции на повреждение ДНК, что приводит к восприятию цитозольной ДНК с помощью пути cGAS/STING (42). Помимо DC, макрофаги могут приобретать TAA, поглощая кальретикулин-положительные клетки. Таким образом, они могут дополнительно способствовать активации CD8 + Т-клеток. Однако этой функции макрофагов противодействует экспрессия SIRPα. Этот рецептор связывается с CD47, который конститутивно экспрессируется при различных типах рака и служит сигналом «не ешь меня» (43).
Другим способом, которым макрофаги могут способствовать элиминации зарождающихся клеток, является высвобождение оксида азота (NO) и TNF-α, которое в экспериментальных условиях может быть вызвано запуском NK2GD (27). Также другие стимулы, более подходящие для человека, такие как IFN-γ, анти-CD40 и лиганды толл-подобного рецептора (TLR), могут индуцировать высвобождение NO и TNF-α, тем самым стимулируя онкоцидную активность макрофагов. (44). По крайней мере, для IFN-γ есть исследования, которые предполагают, что это может происходить на ранних стадиях формирования рака. На модели MHC-II-негативной множественной миеломы было показано, что макрофаги быстро рекрутируются и представляют ТАА CD4 9.0665 + Т-клетки, которые, в свою очередь, активируют программу уничтожения опухолей в макрофагах посредством секреции IFN-γ (45). Также NK-клетки и CD8 + Т-клетки продуцируют высокие уровни IFN-γ, тем самым провоцируя гипотезу о том, что также эти лимфоциты могут стимулировать опухолецидные свойства макрофагов.
Фаза равновесия
Скоординированная иммунная атака, проводимая во время фазы элиминации, может уничтожить трансформированные клетки. Однако продолжающееся давление иммунной системы может привести к отбору раковых клеток, которые могут преобладать. Раковые клетки могут избежать иммунного распознавания несколькими способами, среди которых подавление механизма процессинга антигена или экспрессия молекул TAA или MHC-I. Следовательно, отбираются раковые клетки со сниженной иммуногенностью. Несколько доклинических исследований предоставили доказательства роли CD8 + Т-клеток в этом процессе (46). Клиническая значимость этого была предложена в нескольких исследованиях рака, показывающих избирательное подавление целевого TAA или MHC-I при рецидивирующих опухолях (47–49). Также показано, что миелоидные клетки играют роль в формировании иммуногенности опухолей. Образуя CD8 + Т-клетки, они косвенно способствуют онкогенности. Для TAM1 было предложено более прямое действие (50). Было показано, что в отсутствие Т-клеток NK-клетки могут активировать TAM1 за счет продукции IFN-γ и что эти TAM1 служат модуляторами иммуногенности опухоли. В дополнение к маскировке, раковые клетки кооптируют регуляторные цепи, чтобы ослабить иммунные реакции. К ним относятся, помимо прочего, экспрессия PD-L1, рекрутирование регуляторных Т-клеток (Treg) и регуляторных миелоидных клеток. Многие механизмы, используемые регуляторными клетками для паралича опухолеспецифических эффекторных клеток и, как таковые, для создания толерантной среды для раковых клеток, рассматривались в другом месте (51, 52). Следовательно, опухолевые клоны выдерживают контрольную точку элиминации и образуют клинически скрытую опухоль, которая сосуществует с иммунными клетками хозяина. Во время этого динамического равновесия вблизи опухолевых клеток можно обнаружить иммунные клетки с анти- и проопухолевой активностью (53).
Гибель клеток, совпадающая с присутствием поверхностного кальретикулина и сигналов тревоги, таких как IFN типа I, является ключом к активации противоопухолевого иммунитета DC (41). Этот тип гибели клеток, когда он дополнительно связан с высвобождением аденозинтрифосфата (АТФ), белков группы высокой подвижности 1 (HMGB1) и аннексина A1 (ANXA1), получил название иммуногенной гибели клеток (ICD). В то время как гибель клеток из-за ROS в сочетании со стрессом эндоплазматического ретикулума (ER) приводит к ICD в опухолевых клетках, апоптоз, вызванный воздействием на опухолевые клетки IFN-γ и TNF-α, менее иммуногенен (54). Это может вызвать покой, вовлеченный в спячку опухоли (55, 56). Кроме того, провоспалительные сигналы, которые HMGB1 обычно запускает в миелоидных клетках, сильно уменьшаются, когда этот белок окисляется (57). Более того, длительное производство АФК и АРНК миелоидными клетками может увеличить количество случаев мутации ДНК в опухолевых клетках и, как таковое, само по себе быть причиной прогрессирования рака (58). Несколько механизмов встроены в воспалительный каскад, который происходит во время фазы элиминации, для предотвращения чрезмерного повреждения сопутствующих тканей и аутоиммунитета. Например, выведение дебриса посредством фагоцитоза и аутофагии приводит к секреции TGF-β, который действует как сильный иммунодепрессант и активирует поляризованные TIM 2 типа, что приводит к опосредованному CXCL5 воспалению и росту опухоли (25, 59).). Даже T H 1-связанные цитокины могут способствовать накоплению TIM с регуляторными функциями. Было показано, что как IFN-γ, так и IL-1β участвуют в мобилизации, рекрутировании и ингибирующей активности MDSCs (60). Факторы транскрипции, которые управляют этими событиями, смещаются от провоспалительного IRF8 и преобразователя сигналов и активатора транскрипции (STAT) 1, 2, 3 и 6 к CCAAT-энхансер-связывающему белку β (C/EBPβ) и PI3Kγ, генерируя незрелые TIM. Кроме того, гликолиз, метаболический путь, предпочитаемый ТИМ и опухолевыми клетками во время фазы элиминации, опосредует продукцию белков, нуклеиновых кислот, липидов, лактата, аденозина и, следовательно, иммуносупрессивное закисление ТМЭ. В результате TIM подвергаются гамма-рецептору, активируемому пролифератором пероксисом (PPARγ), и STAT-опосредованному метаболическому перепрограммированию (61). Они переключаются с гликолиза на окислительное фосфорилирование и катаболизм липидов (окисление жирных кислот). Ингибирование пути гликолиза в TAN1, TAM1 и зрелых DC сокращает их активность, тем самым благоприятствуя активности TIM, способствующих развитию опухоли, что в конечном итоге приводит к фазе ускользания, когда опухоли с иммунологической формой постепенно растут, становятся клинически очевидными и имеют установленный иммуносупрессивный TME. (62).
Фаза побега
Ранний иммунный ответ на раковые клетки направлен на устранение опасности и характеризуется острым воспалением. Однако персистенция раковых клеток, совпадающая с сосуществованием иммунных клеток с про- и противоопухолевыми свойствами, приводит к тлеющему воспалению. Это хроническое воспаление в основном поддерживается TAM2, TAN2 и MDSC, которые являются основными подмножествами TIM, обнаруженными во многих установленных раках. Это неудивительно, поскольку опухолевые клетки продуцируют такие факторы, как G-CSF, GM-CSF и VEGF, чтобы обеспечить непрерывный набор миелоидных предшественников в кровоток. В физиологических условиях GM-CSF управляет миелопоэзом, а также дифференцировкой DC, в то время как G-CSF и M-CSF обеспечивают дифференцировку, пролиферацию и выживание гранулоцитов и макрофагов соответственно (10). Однако при хроническом воспалении GM-CSF подавляет IRF8 в предшественниках ДК и, таким образом, приводит к уменьшению развития ДК наряду с привлечением большего количества MDSC. Кроме того, M-CSF подавляет дифференцировку DCs, одновременно усиливая поляризацию TAM2 (63). Поэтому этот фактор считается предиктором плохого прогноза (64). G-CSF может мобилизовать гранулоцитарные миелоидные клетки из костного мозга для стимуляции ангиогенеза. Более того, незрелые ТИМы будут способствовать набору большего количества регулирующих ТИМов, что приведет к «незаживающей ране». Факторами, которые способствуют рекрутированию миелоидных клеток, таких как моноциты, DC, MDSC и TAN2, в TME, среди прочего, являются IL-4, IL-6, IL-8, IL-10, S100A8/A9., VEGF и TGF-β (65).
Опухолестимулирующие функции TIM обычно активируются запуском TLR и связыванием цитокинов и факторов роста, таких как IFN-γ, IL-4, IL-6, IL-10, IL-13 и TGF-β, в ответ на их реакцию. рецептор. Сигнальный каскад после взаимодействия с рецептором приводит к усилению различных факторов транскрипции, включая STAT1, 3 и 6, митоген-активируемую протеинкиназу, регулируемую внеклеточно (MAPK-ERK) и ядерный фактор-κB (NF-κB), при этом STAT3 является ведущим. регулятор в большинстве субпопуляций миелоидных клеток (66-68). После активации TIM выполняют перекрывающиеся функции. TAM2 и MDSC хорошо известны своей способностью подавлять опосредованное Т-клетками уничтожение опухоли (рис. 3), поскольку оба могут оказывать прямое иммуносупрессивное действие за счет межклеточного контакта (например, CD80 и PD-L1), секретируемых факторов (например, IL-1). 10 и TGF-β) и экспрессию ферментов (например, ARG-1, iNOS, IDO) (69–75). Кроме того, TAM2 и MDSC задействуют Tregs, чтобы помочь им подавить противоопухолевый иммунный ответ. С этой целью эти TIM экспрессируют и продуцируют несколько факторов, которые стимулируют дифференцировку (например, IDO, CD40, простагландин E2 [PGE2], IL-10, TGF-β) или рекрутирование Treg (например, CCL22) (76–78). ). TAM2 может дополнительно вмешиваться в активность TAM1, подавляя экспрессию IFN-γ и IL-12, тем самым влияя на прямое уничтожение опухолевых клеток, а также на активацию Т-клеток-киллеров (65). Хотя TAN2 не известны своей подавляющей Т-клетки активностью, было показано, что устранение TAN2 приводит к повышению активности цитотоксических Т-клеток, явление, которое было связано с TGF-β, который способствует накоплению TAN2 и индуцирует экспрессию ARG-1 (16, 79). Хотя ДК не присутствуют в большом количестве в установленном TME, важно обсудить их роль в облегчении побега опухолевых клеток от иммунного киллинга (рис. 3). Условия в TME, такие как накопление внеклеточного аденозина и лактата, VEGF, M-CSF, TGF-β, IL-6 и гипоксия, совместно ингибируют презентацию антигена DCs, тем самым ингибируя активацию адаптивного противоопухолевого иммунитета (80–82). ). Более того, во время опухолевой прогрессии субпопуляции ДК способны достигать дренирующих опухоль лимфатических узлов и стимулировать пролиферацию Treg зависимым от TGF-β образом, превращая незрелые ДК из жертвы в соучастников преступления (83). Было даже описано, что зрелые DC внутри TME могут проявлять проопухолевые свойства посредством экспрессии IDO, ARG-1 и iNOS. Эти зрелые, но tolDC блокируют пролиферацию Т-клеток и в присутствии TGF-β дополнительно стимулируют дифференцировку Treg (84-87).
Стимулирование ангиогенеза за счет секреции VEGF, ангиопоэтина 1 и 2 и GM-CSF, а также диссеминация опухолевых клеток за счет секреции MMP и катепсинов — две другие общие черты TAN2, tolDC, TAM2 и MDSC (88). Более того, было высказано предположение, что TIM, такие как MDSCs и TAN2, ответственны за создание преметастатической ниши (89–93). В этих предметастатических нишах вырабатываются факторы для привлечения и последующей остановки опухолевых клеток, такие как IL-1β, прокинетин 2 и ММП, в то время как соседние клетки стимулируются к секреции VEGF для облегчения прибытия опухолевых клеток.
В заключение, TME инструктирует TIM выполнять функции в соответствии с потребностями опухоли. Эти функции связаны с хроническим воспалением, ремоделированием тканей и иммуносупрессией и способствуют росту опухоли и метастазированию, поскольку эти функции позволяют опухолевым клеткам получать питательные вещества, пролиферировать, выживать, мигрировать и избегать элиминации иммунной системой.
Терапия рака и миелоидные клетки: взаимное влияние и последствия
Экспериментальные исследования предоставили достаточно доказательств того, что миелоидные клетки влияют практически на все виды терапии рака, начиная от химиотерапии (КТ), лучевой терапии (ЛТ), таргетной терапии опухоли и заканчивая иммунотерапией (рис. 4). Поэтому в следующих разделах будет кратко обсуждено взаимодействие с различными методами лечения рака.
Рисунок 4 . Двойная роль миелоидных клеток в ответах на противораковую терапию. (A) Схемы ЛТ и КТ могут индуцировать ИКД, тем самым предупреждая антигенпрезентирующие клетки (АПК) об опасности, которую представляют раковые клетки. Эта тревога подается за счет высвобождения различных DAMP (например, HMGB1, АТФ и т. д.) и облегчает перекрестное примирование опухолеспецифических CD8 + Т-клеток в дренирующих опухоль лимфатических узлах. Активированные Т-клетки CD8 + впоследствии проникают в опухоль в поисках раковых клеток, экспрессирующих ТАА, и после распознавания используют различные механизмы для уничтожения раковых клеток. В то время как TAM1 и TAN могут дополнительно повышать эффективность CT за счет высвобождения ROS, тем самым усиливая гибель опухолевых клеток, другие TIM, включая MDSC, могут противодействовать эффектам RT и CT не в последнюю очередь за счет ингибирования CD8 + Т-клетки. (B) Внутри TME CD8 + Т-клетки часто становятся толерантными с помощью иммуносупрессивных факторов, экспрессируемых tolDC, TAM2, MDSC, опухолевыми клетками (и Treg). Одной из стратегий является экспрессия PD-L1, который связывается с PD-1 на активированных Т-клетках. Следовательно, было изучено ингибирование иммунных контрольных точек для облегчения опосредованной PD-L1: PD-1 иммуносупрессии. Моноклональные антитела активно используются для этой цели, однако было показано, что они захватываются ТАМ с использованием их FcγR и противодействуют MDSC. Тем не менее успех ингибирования иммунных контрольных точек коррелирует с наличием иммуностимулирующих TIM и CD8 9.0665 + Т-клеток в пределах TME. (C) Было показано, что низкомолекулярные ингибиторы, нацеленные на протеинкиназы, участвующие в развитии опухолевых клеток, оказывают влияние на TIM. Некоторые из этих эффектов полезны для исхода терапии. Например, ингибиторы BRAF обращают вспять подавление, оказываемое клетками меланомы на TADC. Однако большинство этих эффектов потенцируют ТИМ для выполнения функций, способствующих развитию опухоли, или предотвращают попадание ТИМ в ТМО, чтобы противостоять росту опухоли. Примерами здесь являются иматиниб, который стимулирует поляризацию TAM1 к TAM2, ингибиторы BRAF, которые восстанавливают компартмент MDSC посредством индукции CCL2, и ингибиторы MET, которые предотвращают рекрутирование нейтрофилов с цитотоксической способностью (iNOS-опосредованное высвобождение NO).
Миелоидные клетки и лучевая терапия
Лучевая терапия (ЛТ) использует ионизирующее излучение для уничтожения злокачественных клеток и может индуцировать полноценный ИКД дозозависимым образом (94). ICD позволяет TADC приобретать TAA, а также стимулы, которые усиливают их созревание, что позволяет им активировать опухолеспецифические CD8 + T-клетки (95, 96). Более того, низкие дозы ЛТ могут перепрограммировать ТАМ в ТАМ1, что может способствовать рекрутированию и активации опухолеспецифических Т-клеток (97). Однако иммуногенные эффекты, опосредованные лучевой терапией, могут быть замаскированы (9).8, 99). В установленном TME TIM инструктируют противодействовать противоопухолевым иммунным ответам. В зависимости от дозы, ее фракционирования, а также модели опухоли РТ может рекрутировать миелоидные клетки и даже поляризовать их в иммуносупрессивные клетки, способствующие развитию опухоли (100). Последний вывод предполагает, что истощение TIM может быть стратегией улучшения результатов ЛТ. Действительно, в нескольких экспериментальных исследованиях было показано, что истощение ТАМ, MDSC и нейтрофилов повышает эффективность ЛТ (101–103). Однако следует соблюдать осторожность, чтобы гарантировать, что опосредованные лучевой терапией эффекты на TADC и поляризацию TAM1 не будут потеряны, поскольку оба они способствуют абскопальным эффектам лучевой терапии.
Миелоидные клетки и химиотерапия
Химиотерапия (КТ) бывает разных цветов и форм. Как и в случае с лучевой терапией, некоторые режимы КТ, такие как использование антрациклинов, могут индуцировать полноценный ICD, тем самым воздействуя на TADC, которые, в свою очередь, активируют опухолеспецифические CD8 + T-клетки. Было показано, что эффективность антрациклинов зависит от TADC (104, 105). Также ТАМ могут играть полезную роль во время лечения КТ. Эти клетки могут действовать как депо лекарств, когда соединения CT доставляются в виде наночастиц, тем самым обеспечивая местную и пролонгированную доставку терапии (106). Кроме того, при лечении оксалиплатином наблюдается продукция АФК ТАМ и ТАН, что помогает стимулировать гибель опухолевых клеток (107). Химиотерапия может дополнительно воздействовать на числа TIM, либо истощая, либо усиливая их. Гемцитабин и 5-фторурацил являются примерами химиотерапии, снижающей количество MDSC. Ослабляя иммуносупрессию, опосредованную MDSC, эти методы лечения активируют CD8 + Т-клеточно-зависимые противораковые иммунные ответы (108). Другие химиотерапевтические препараты, такие как паклитаксел, имеют более сложный профиль. В то время как паклитаксел истощает MDSC (которые экспрессируют редуктазу Р450), он увеличивает ТАМ (12, 109). В последнем случае ингибирование накопления ТАМ улучшало результаты терапии (110). Это исследование предполагает, что ТАМ могут снижать эффективность КТ. Это подтверждается несколькими другими исследованиями и, к сожалению, не ограничивается ТАМ. Также MDSC были связаны с отказом CT. ТАМ и MDSC могут индуцировать лекарственную устойчивость, например, путем продукции цистеиновых катепсинов, которые защищают опухолевые клетки от уничтожения химиотерапевтическими агентами, такими как таксол, и даже способствуют росту опухоли, усиливая хроническое воспаление (111, 112). Более того, ТАМ могут секретировать IL-10 в ответ на ХТ и тем самым усиливать иммуносупрессию (113). Таким образом, ТИМы являются одновременно и друзьями, и врагами при лечении онкологических больных с помощью химиотерапии (и ЛТ), что ставит задачу идентифицировать и избирательно воздействовать на врага, оставляя дружественные ТИМ невредимыми.
Миелоидные клетки и опухоли, нацеленные на низкомолекулярную терапию рака
Наши растущие знания о драйверных мутациях, которые обеспечивают раковым клеткам преимущество в росте и выживании, позволили разработать низкомолекулярные препараты, нацеленные на молекулярные изменения, возникающие в результате этих мутаций. Белки с киназной активностью, такие как рецептор фактора стволовых клеток (c-kit), BRAF, MEK и MET, являются примерами мишеней, для которых были клинически протестированы низкомолекулярные ингибиторы. В клинических испытаниях эти низкомолекулярные ингибиторы первоначально уменьшали размер опухоли (114, 115). Однако они мало влияют на долгосрочную перспективу, что может быть, по крайней мере частично, связано с активностью миелоидных клеток. Например, иматиниб, ингибитор тирозинкиназы, который действует посредством активации мутаций c-kit, был оценен при раке желудочно-кишечного тракта. Было показано, что иматиниб индуцирует апоптоз опухолевых клеток. Хотя это предполагаемый результат, далее наблюдалось, что TAM1, который в основном заселял строму желудочно-кишечного тракта, был поляризован на TAM2 из-за активации C/EBPβ при взаимодействии с умирающими опухолевыми клетками (116). BRAF и MEK представляют собой протеинкиназы пути MAPK-ERK, которые в солидных опухолях, таких как меланома, обеспечивают пролиферацию и выживание клеток. Следовательно, мутации BRAF и MEK, которые постоянно активируют MAPK-ERK, связаны с онкогенезом. Для противодействия неконтролируемому росту и выживанию раковых клеток из-за мутаций BRAF и MEK были разработаны небольшие лекарственные ингибиторы (117–120). Развитие резистентности к ингибиторам BRAF в модели меланомы мыши было связано с восстановлением компартмента MDSC. Это зависело от реактивации пути MAPK с последующей продукцией CCL2, аттрактанта миелоидных клеток. Модулирование MDSC с использованием истощающих антител (анти-Gr-1) или блокирование их рекрутирования (антагонист CCR2) препятствовало росту этих устойчивых к BRAF опухолей меланомы (121). Также ТАМ и полученный из ТАМ TNF-α были вовлечены в неэффективность ингибиторов BRAF в моделях меланомы у мышей (122). Блокирование развития ТАМ (ингибирование M-CSFR) повышало эффективность ингибиторов BRAF (123). С другой стороны, ингибиторы MEK и BRAF восстанавливают подавление TADC, вызванное мутировавшими клетками меланомы in vitro (124, 125). Наконец, МЕТ (рецептор фактора роста гепатоцитов) считается молекулярной мишенью при некоторых видах рака, включая немелкоклеточный рак легкого, рак желудочно-кишечного тракта и гепатоцеллюлярную карциному (114). Ингибирование MET также влияет на нейтрофилы, которые полагаются на активность MET, чтобы добраться до опухолей и вызвать цитотоксические реакции, такие как iNOS-опосредованное высвобождение NO (126). Приведенные выше исследования показывают, что необходимы дальнейшие исследования, чтобы понять, как ингибирующие киназы влияют на миелоидные клетки и как можно манипулировать миелоидными клетками, чтобы обеспечить более устойчивый ответ у пациентов.
Миелоидные клетки и иммунная контрольная терапия
Иммунная активация и активность жестко регулируются несколькими механизмами во избежание иммунопатологии (например, аутоиммунитета). Раковые клетки захватывают эти механизмы, чтобы избежать иммунного надзора. Они экспрессируют лиганды, такие как PD-L1, который запускает PD-1, ингибирующий рецептор, экспрессируемый на опухоле-реактивных иммунных клетках, таких как CD8 + Т-клетки. Следовательно, опухоле-реактивные иммунные клетки становятся парализованными. Моноклональные антитела (мАт), которые связывают PD-1 или PD-L1 и блокируют эту иммунную контрольную точку, открывают новую эру в иммунотерапии рака (127–129).). На сегодняшний день пять mAb, нацеленных на путь PD-1:PD-L1, были одобрены для лечения пациентов с солидными опухолями, включая меланому и немелкоклеточный рак легкого. Эти mAb представляют собой пембролизумаб и ниволумаб, которые противодействуют PD-1, и авелумаб, дарвулумаб и атезолизумаб, которые противодействуют PD-L1. Эти mAb стали поворотным моментом в лечении значительного числа пациентов (130). К сожалению, весь потенциал этих mAb еще не реализован.
Значительное число пациентов и раковых заболеваний не поддаются лечению в начале лечения или становятся устойчивыми в ходе курса лечения и, следовательно, терпят неудачу. Существует несколько гипотез, объясняющих эту неудачу терапии, одна из них заключается в том, что TIM ответственны за устойчивость к терапии, блокирующей иммунные контрольные точки. Эта гипотеза возникла в результате наблюдения, что высокая инфильтрация опухолей иммуносупрессивными миелоидными клетками коррелирует с плохим прогнозом и резистентностью к терапии иммунологическими контрольными точками (131, 132). В основном MDSC и ТАМ вовлечены в устойчивость к mAb, блокирующим иммунные контрольные точки. Следовательно, простого блокирования взаимодействий PD-1:PD-L1 может быть недостаточно для преодоления TIM-опосредованного ингибирования CD8 9.0665 + Т-клетки. Это направление мысли подкрепляется наблюдением, что истощение MDSC в экспериментальных моделях, как было показано, усиливает противоопухолевый иммунный ответ и преодолевает резистентность (133). Более того, функциональная модуляция MDSC эпигенетическими препаратами повысила чувствительность нескольких экспериментальных моделей рака, которые ранее были устойчивы к терапии иммунными контрольными точками (134, 135). Описано, что другие ТАМ ингибируют терапию PD-1:PD-L1 путем удаления моноклональных антител против PD-1 из PD-1 + CD8 + Т-клеток, использующих их FcγR (136). Кроме того, новые данные сообщают, что как метаболические, так и воспалительные пути могут усиливать экспрессию PD-L1 на миелоидных клетках (71, 137, 138). Эти исследования предоставляют первое доказательство того, что иммуносупрессивные TIM могут препятствовать терапии иммунной контрольной точки с помощью нескольких избыточных стратегий. Однако есть также доказательства того, что иммуностимулирующие ТИМ являются необходимым условием для успеха иммунных контрольных точек. Было показано, что связывание моноцитов и ДК с помощью mAb против PD-L1, по крайней мере, частично отвечает за последующий контроль роста опухоли (139)., 140). Потребность в ДК для гарантии успешной терапии контрольных точек была дополнительно подтверждена в экспериментах, в которых доставка ДК, происходящих из костного мозга, к опухолям приводила к рекрутированию Т-клеток и сенсибилизации к анти-PD-1 терапии (141). Подобно заключению о миелоидных клетках в ЛТ, КТ и таргетной терапии, мы можем заключить, что иммунная контрольная точка может поддерживаться союзниками ТИМ, т. CD8 + Т-клетки, можно рассматривать как саботажников этой терапии. Сообщалось, что помимо пути PD-1:PD-L1 другие рецепторы, ингибирующие контрольные точки, или их лиганды экспрессируются одним или несколькими подмножествами TIM. Например, экспрессия Т-клеточного иммуноглобулина и муцинового домена, содержащего-3 (TIM-3), была описана в TADC, в то время как «V-домен Ig-супрессор активации Т-клеток» (VISTA) преимущественно экспрессируется на гемопоэтических клетках, и в нескольких моделях рака мышей обнаружены особенно высокие уровни TIM. Кроме того, сообщалось, что «Т-клеточный иммунорецептор с доменами Ig и ITIM» (TIGIT) связывает CD155 на DC и макрофагах, в то время как основной лиганд «гена активации лимфоцитов 3» (LAG-3) представлен MHC-II, экспрессируемым в разной степени практически на всех ТИМах. Поэтому необходимы дальнейшие исследования, чтобы выяснить, какую роль TIM играют в других механизмах ингибирования контрольных точек.
Стратегии, разработанные для нацеливания на миелоидные клетки с целью лечения рака
Публикуется все больше экспериментальных и клинических исследований по нацеливанию на миелоидные клетки при раке, при этом за последние 5 лет было опубликовано около 400 исследований. В целом подходы к нацеливанию миелоидных клеток с характеристиками, способствующими развитию опухоли, такими как TAM2, TAN2 и MDSC, можно разделить на две основные стратегии; сокращение их количества путем их истощения или блокирования их рекрутирования и реполяризации их функций. И наоборот, были предприняты попытки увеличить количество и функцию миелоидных клеток с противоопухолевыми свойствами. В следующих разделах мы выделяем несколько многообещающих стратегий (рис. 5).
Рисунок 5 . Смещение баланса в сторону миелоидных клеток с противоопухолевым фенотипом. Было изучено несколько подходов для увеличения соотношения анти- и проопухолевых ТИМ. К ним относятся истощение или реполяризация ТИМ, способствующих развитию опухоли, а также привлечение и активация противоопухолевых ТИМ.
Манипулирование номерами миелоидных клеток
Истощение миелоидных клеток и даже специфических подмножеств в качестве стратегии уменьшения числа миелоидных клеток может осуществляться с использованием антител, нацеленных на специфические поверхностные продуценты миелоидных клеток, такие как CD11b, Gr-1 или Ly6G. Кроме того, линии трансгенных мышей также можно использовать с постоянным или условным удалением миелоидных клеток. Условная делеция позволяет избежать эмбриональной летальности, в то же время препятствуя анализу функции гена в четко определенный момент времени во взрослых тканях. Две наиболее часто используемые модели основаны на добавлении дифтерийного токсина и/или рекомбиназы Cre/LoxP. Первая модель основана на нокауте рецептора дифтерийного токсина позади промотора маркера, характеризующего тип клеток, такого как CD11b или CD11c-рецептор дифтерийного токсина у мышей. Напротив, получение мышей с условно нокаутным Cre/LoxP представляет собой многостадийный процесс, который включает скрещивание мутанта flox с тканеспецифичными Cre-экспрессирующими линиями мышей. Таким образом, последний позволяет проводить условную специфическую абляцию подтипа миелоидных клеток в конкретной интересующей ткани.
Такие экспериментальные подходы доказывают, что истощение миелоидных клеток с иммуносупрессивными функциями может задерживать рост опухоли, в то время как истощение стимулирующих миелоидных клеток имеет противоположный эффект (142, 143). Поскольку эти подходы не ограничивают истощение миелоидных клеток ТМЭ, тем не менее, нет уверенности в том, что наблюдаемые эффекты могут быть частично приписаны истощению миелоидных клеток на периферии. Более того, стратегии полного истощения непривлекательны для применения в условиях человека. Поэтому были разработаны другие фармакологические и основанные на генах подходы, чтобы препятствовать накоплению супрессивных миелоидных клеток. Они основаны на наших текущих знаниях о том, как миелопоэз и рекрутирование TAM, MDSC и TAN управляются. Накопление ТАМ в ТМО сильно зависит от передачи сигналов CCL2:CCR2 и M-CSF:M-CSFR, поэтому существуют разные подходы с использованием моноклональных антител (144–146), низкомолекулярных ингибиторов (145, 147, 148) и РНК-интерференции (149).) были разработаны, чтобы блокировать эти пути. В целом эти исследования показали снижение числа ТАМ и коррелирующее с этим замедление роста опухоли в различных моделях рака у мышей. Эти исследования также предоставили информацию о влиянии блокады CCL2:CCR2 и M-CSF:M-CSFR, а также о том, как схемы лечения могут вызывать нежелательные эффекты.
При прекращении блокады оси CCL2:CCR2 у мышей наблюдался внезапный выброс моноцитов из костного мозга, следствием чего было увеличение частоты метастазирования и связанное с этим снижение выживаемости животных (110). Использование мышей с нокаутом CCL2 по сравнению с мышами с нокаутом CCR2 также показало, что дефицит CCL2 приводит к задержке роста карциномы молочной железы, в то время как дефицит CCR2 усиливает рост опухоли (150). Различия в программах транскрипции в моноцитах с дефицитом CCR2 по сравнению с CCL2 могут объяснить эти несопоставимые эффекты нарушения CCR2 по сравнению с CCL2 на прогрессирование карциномы молочной железы. Хотя это исследование не показало различий в инфильтрации моноцитов или ТАМ, возможно, что другие подмножества ТИМ могут играть роль. Например, MDSC также могут воздействовать на CCL2 посредством экспрессии CXCR2 (151). Подобно вмешательству в ось CCL2:CCR2 и накоплению ТАМ, влиянию как на CCR2 + TAM и CXCR2 + TAN оказывают сильное воздействие, по крайней мере, при использовании в сочетании с КТ (152, 153). Поэтому крайне важно определить всех соответствующих игроков при блокировании определенной молекулы, чтобы полностью понять потенциальные сильные и слабые стороны подхода истощения.
Воздействие на путь M-CSF:M-CSFR также препятствует накоплению ТАМ. Этот подход задерживает рост опухоли во всех протестированных экспериментальных моделях либо при использовании в качестве самостоятельной терапии (146, 147), либо при использовании в сочетании с другими методами лечения (101, 102, 109)., 113). В некоторых исследованиях, в которых был нацелен путь M-CSF:M-CSFR, сообщалось о регрессии опухоли, которая не была связана с количественными изменениями ТАМ в TME. Однако в этих исследованиях сообщалось об изменениях в фенотипе ТАМ, особенно о подавлении связанных с ТАМ2 признаков (145, 149). Поскольку прерывание передачи сигналов CCL2:CCR2 и M-CSF:M-CSFR было полезным во многих экспериментальных моделях, эти пути оцениваются у пациентов как при монотерапии (146), так и при комбинированной терапии (КТ и терапия, блокирующая иммунные контрольные точки) (154). .
Другая стратегия, которая изучается в контексте уменьшения количества миелоидных клеток, способствующих развитию опухоли, в TME заключается во вмешательстве в передачу сигналов VEGF:VEGFR. VEGFR в высокой степени экспрессируется TIM, и взаимодействие с VEGF способствует их миграции и дифференцировке в иммуносупрессивные клетки (155). Таким образом, вмешательство в передачу сигналов VEGF:VEGFR может играть двойную роль в отношении TIM, снижая их количество и реполяризуя их функцию (в дополнение к влиянию на ангиогенез). Было показано, что с точки зрения привлечения TIM передача сигналов VEGF:VEGFR привлекает макрофаги (156) и моноциты зависимым от CCR2 образом (157). Кроме того, было показано, что рекрутирование MDSCs в TME немелкоклеточной карциномы легкого снижается при блокаде передачи сигналов VEGF:VEGFR. Однако в этом исследовании привлечение моноцитов увеличивалось при лечении антителами против VEGF (158). Несколько препаратов на основе анти-VEGF, включая бевацизумаб, авастин и акситиниб, нацеленные на VEGFR, используются для лечения различных видов рака человека (159). –161). В то время как препараты, направленные против VEGFR, используемые в этих исследованиях, в основном изучаются с точки зрения ангиогенеза, было показано, по крайней мере, для препарата акситиниба, что он воздействует на миелоидные клетки. В ДК, подвергшихся воздействию акситиниба, наблюдалось дозозависимое снижение активации STAT3. Несмотря на это благоприятное изменение, у этих ДК была нарушена их стимулирующая активность Т-клеток (162). Напротив, на модели метастазирования меланомы в головной мозг было показано, что акситиниб увеличивает количество M-MDSC и меняет их функцию с подавления Т-клеток на стимулирование клеток с последующей задержкой прогрессирования заболевания (163). Таким образом, это исследование предполагает, что обогащение миелоидных клеток противоопухолевыми свойствами может быть привлекательным подходом к лечению рака. Эта идея подтверждается экспериментальными и клиническими исследованиями, в которых ex vivo ДК (часто сконструированные для секреции стимулирующих цитокинов), доставленные в опухоль, опосредуют контроль заболевания (164, 165).
Использование Flt3-L для увеличения количества DC широко изучалось, т.к. Flt3:Flt3-L опосредованная передача сигналов участвует в генерации различных субнаборов DC из клеток-предшественников костного мозга (166, 167). В экспериментальных моделях введение Flt3-L (самостоятельно или в сочетании с ХТ) увеличивало количество ДК и приводило к задержке роста опухоли (168–171). Доказательства, подтверждающие использование Flt3-L в клинических условиях, были получены в исследовании, в котором рекомбинантный человеческий Flt3-L вводили для увеличения числа DC 9.0461 in vivo . Эти ДК собирали с помощью лейкафереза, импульсно вводили пептиды, полученные из опухолевого антигена, и использовали для вакцинации, вызывая регрессию опухоли у 2 из 12 пациентов (172).
Также GM-CSF изучался как средство для увеличения количества DC in vivo . Это может показаться спорным, учитывая роль GM-CSF в миелопоэзе. Это, безусловно, стратегия, к которой следует подходить с осторожностью, поскольку было показано, что блокирование GM-CSF снижает накопление моноцитов и миелоидных клеток-предшественников и задерживает прогрессирование опухоли при доставке GM-CSF к опухолям, например, с использованием было показано, что онколитический вирус T-VEC индуцирует опухолеспецифические Т-клетки, скорее всего, в результате активации DC. В этом клиническом исследовании доставка GM-CSF с помощью T-VEC также привела к уменьшению количества MDSC (173, 174). Последнее удивительно, поскольку конструирование опухолей для экспрессии высоких уровней GM-CSF было связано с накоплением MDSC в различных экспериментальных моделях (175, 176).
В целом эти исследования подтверждают идею о том, что уменьшение количества TIM с активностью, способствующей развитию опухоли, при одновременном увеличении количества TIM с противоопухолевой активностью является ценным подходом в борьбе с раком. Однако эти исследования также указывают на то, что смещение баланса в сторону увеличения количества противоопухолевых TIM за счет воздействия на пути, участвующие в миелопоэзе, является сложной задачей.
Управление функцией миелоидных клеток
Миелоидные клетки характеризуются высокой пластичностью. Их способность изменять форму в соответствии с сигналами, которые они получают, инициировала исследования стратегий, которые способствуют их анти- и / или ингибированию их проопухолевой активности.
Управление генетическими программами в миелоидных клетках
Генетические программы, контролируемые транскрипционными сетями и эпигенетическими механизмами, определяют фенотип и функции миелоидных клеток. Поэтому их перепрограммирование было в центре внимания. Запуск TLR на TIM, в частности TLR3 и TLR9, был широко изучен в этом контексте, поскольку эти сети запускают транскрипцию. Показано, что Т-клеточно-супрессорная активность MDSC снижается при воздействии агонистов TLR3 или TLR9.(177–180). Воздействие на MDSC лиганда TLR9 CpG стимулировало их выработку T H 1, активирующих цитокины, и даже дифференцировку в TAM1, переключатель, управляемый IFN-α, продуцируемый плазмоцитоидными DCs после стимуляции CpG (178, 181). Помимо своего влияния на MDSCs, запуск TLR3 также превращает TAM2 в TAM1 и облегчает перекрестное примирование антиген-специфических CD8 + T-клеток с помощью TADC, тем самым способствуя регрессии опухоли (182, 183).
Использование TLR4 для функционального перепрограммирования DC было достигнуто с помощью уникального генетического подхода DC, что, вероятно, объясняет его успех, поскольку ранее было показано, что MDSC, инициированные воспалением, активируются липополисахаридом (LPS), хорошо известным лигандом TLR4. , чтобы производить повышенные уровни IL-10 и подавлять продукцию IL-12 макрофагами (184).
Взаимодействие CD40:CD40-лиганд (CD40L) может представлять собой еще одну интересную мишень для подавления проопухолевой программы транскрипции MDSCs, одновременно стимулируя противоопухолевую программу транскрипции TAM и TADC. Было показано, что mAb-агонисты против CD40 блокируют связь MDSC:Treg и способствуют созреванию MDSC (185, 186). Макрофаги активируются при лечении этими моноклональными антителами против CD40, что позволяет им проникать в опухоли и оказывать противоопухолевое действие (187). Наблюдение, что mAb против CD40 активируют CD8 + Независимая от Т-клеток гибель опухолевых клеток побудила к использованию полностью гуманизированных моноклональных антител против CD40 (CP-870.893) в фазе I клинических испытаний, показавших, что их комбинация с гемцитабином активирует противоопухолевый иммунный ответ (188). Активация DC с помощью CD40 позволяет им выполнять свою функцию стимуляции CD8 + Т-клеток (189). Было показано, что сочетание лигандов CD40 и TLR3 наделяет tolDC человека и мыши стимулирующими свойствами (190). Также изучали TLR4 в дополнение к доставке CD70 в сочетании с CD40-активацией TADCs (191). Было показано, что TADC, модифицированные CD70 и одновременно активированные через CD40 и TLR4, способны мигрировать в дренирующие опухоль лимфатические узлы и стимулировать опухолеспецифические CD8 + Т-клетки.
Цитокины представляют собой еще одно средство функционального перепрограммирования миелоидных клеток. В этом контексте было показано, что IL-12 и IFN типа I стимулируют противоопухолевые свойства, в то время как IL-10 и TGF-β связаны со свойствами, способствующими развитию опухоли. Следовательно, было изучено воздействие на TIM IL-12, IFN-α или IFN-β и/или их экранирование от IL-10 и TGF-β. Было показано, что доставка IL-12 в TME оказывает существенное противоопухолевое действие, которое коррелирует с перепрограммированием MDSC, TADC и TAM в антигенпрезентирующие клетки с CD8 9. 0665 + Активирующая способность Т-клеток (175, 192–196). Противоопухолевые стимулирующие свойства ИЛ-12 были подтверждены клиническими реакциями, наблюдаемыми у пациентов с почечно-клеточным раком, меланомой и перитонеальными метастазами рака яичников при лечении ИЛ-12 (197–202). IFN типа I, в частности IFN-α и IFN-β, давно показали, что они оказывают онкоцидное действие и действуют как сильные индукторы поляризации миелоидных клеток, в основном для MDSC и TADC (203, 204). Хотя IFN-α показал многообещающие клинические результаты при гематопоэтическом раке, при солидных опухолях был достигнут небольшой успех (205). Это может быть связано с его системным введением или использованием в качестве монотерапии в этих исследованиях. В доклинических исследованиях было показано, что сочетание IFN-β-стимуляции миелоидных клеток с одновременной блокадой передачи сигналов TGF-β является жизнеспособной стратегией для перепрограммирования MDSC и DC, тем самым облегчая CD8 + ответов Т-клеток (206). Также IL-10 включает иммуносупрессивные программы транскрипции в миелоидных клетках. Антитела, блокирующие рецептор IL-10, используемые отдельно или в сочетании с CpG, восстанавливали экспрессию IL-12 посредством активации DC и стимулировали противоопухолевый ответ (113). Однако в некоторых исследованиях удаление IL-10 было связано с повышенным уровнем MDSCs и Treg в TME и в дренирующих опухоль лимфатических узлах (207).
Также были разработаны стратегии более прямого управления программами транскрипции в миелоидных клетках, включая подходы к подавлению или усилению транскрипционных факторов, таких как STAT3 и NF-kB (208, 209).). Фактор транскрипции STAT3 считается главным регулятором иммуносупрессивной активности миелоидных клеток (66, 67). Исследования низкомолекулярных ингибиторов, действующих на передачу сигналов JAK2:STAT3, показали задержку роста опухоли, которая в некоторых случаях совпадала с переключением TAM2 на TAM1, увеличением MDSCs или не зависела от модуляции TIM (210, 211). В клинических испытаниях низкомолекулярные ингибиторы показали ограниченную эффективность и значительные побочные эффекты, что побудило к разработке более целенаправленных и более конкретных стратегий, основанных на РНК-интерференции и олигонуклеотидах-ловушках. Некоторые из них прошли клинические испытания (212–214). Более селективная доставка STAT3, ингибирующих малую интерферирующую РНК (siRNA), была достигнута путем их связывания с CpG. Следовательно, эта стратегия использует положительное влияние CpG на TLR9.положительные миелоидные клетки, а также потенциальные эффекты ингибирования STAT3. После лечения TLR9, экспрессирующие миелоидные клетки, такие как PMN-MDSC, показали снижение иммуносупрессивной способности. Также опухолевые клетки, экспрессирующие TLR9, теряли устойчивость к апоптозу, вмешиваясь в передачу сигналов STAT3 (212, 213, 215, 216). Менее активированный статус TADC и их общая сниженная способность реагировать на стимуляцию TLR были связаны с гиперактивностью STAT3 (215). Поэтому ингибирование STAT3 было изучено в контексте модуляции TADC. Здесь была использована способность TADC поглощать наночастицы для доставки миРНК STAT3 и полиинозиновой: полицитидиловой кислоты (polyI:C), хорошо известного агониста TLR3. Эта стратегия позволила TADC повышать экспрессию костимулирующих молекул и IL-12 и индуцировать мощные противоопухолевые иммунные ответы (215). Известно, что помимо STAT3, NF-kB регулирует экспрессию многих генов, способствующих развитию опухоли, включая VEGF, IL-6, TNF-α и циклооксигеназу 2 (COX2), что подтверждает его решающую роль в активации TAM2 в ТМЭ (217, 218). Ингибирование NF-κB позволяет ТАМ приобретать тумороцидный фенотип, что приводит к in vivo при регрессии запущенных опухолей (219). В TADC активность NF-κB ингибируется FOXO3, что связано с иммуносупрессивной активностью TADC (220). Активация NF-κB в ДК широко изучалась, хотя в основном в контексте противораковой вакцинации. В целом эти исследования показали, что селективная активация NF-κB в ДК человеческого или мышиного происхождения повышает их способность активировать CD8 + Т-клетки (209, 221). Другим фактором транскрипции, который недавно привлек внимание, является PI3Kγ, поскольку было показано, что этот фактор транскрипции регулирует экспрессию ARG-1 и TGF-β в ТАМ, поэтому представляет собой еще один важный регулятор иммуносупрессивного фенотипа ТАМ (222). Более того, PI3Kγ в миелоидных клетках был напрямую связан с успехом терапии иммунных контрольных точек, указывая на то, что PI3Kγ является привлекательной мишенью для иммунотерапии рака (222, 223).
Генетические программы, активные в миелоидных клетках, также регулируются на эпигенетическом уровне (138). Альтернативно активированные макрофаги костного мозга имеют повышенный уровень ацетилированных гистонов. Это было связано с экспрессией генов TAM2 (224). Бромодоменные белки и белки с экстратерминальным мотивом (225) или ингибиторы гистондеацетилазы (HDACi) (226–228), которые препятствуют ремоделированию хроматина, склонны к фенотипу TAM1 и, как было показано, опосредуют регрессию опухоли в ответ на иммунную контрольную терапию в модели в которых монотерапия антителами против PD1 оказалась неэффективной (228). Также было показано, что HDACi численно и функционально влияет на MDSC (134, 135). Ингибирование HDAC снижало количество MDSC, а также экспрессию характерных ферментов, включая ARG-1 и iNOS, что делало несколько экспериментальных моделей рака чувствительными к терапии иммунных контрольных точек (134, 135). HDACi в настоящее время изучается в комбинации с блокаторами контрольных точек у пациентов с метастатическим и нерезектабельным HER2/neu-отрицательным раком молочной железы (138).
Манипулирование метаболизмом миелоидных клеток
Поскольку метаболизм ТИМов связан с их функцией, изменение метаболических путей изучалось как стратегия воздействия на миелоидные клетки (229). MDSC имеют повышенную экспрессию CD36, транслоказы жирных кислот, которая обеспечивает поглощение и окисление жирных кислот и, таким образом, увеличение их иммуносупрессивной способности (230). Ингибирование окисления жирных кислот снижает иммуносупрессивную способность MDSCs и, кроме того, опосредует противоопухолевый эффект в экспериментальных моделях в сочетании с низкодозовой ХТ и адоптивной клеточной терапией (231). Метаболические пути также изучались в TADC. Метаболизм жирных кислот в ДК был связан с их способностью перекрестно представлять антигены, критический процесс для стимуляции опухолеспецифических CD8 9.0665 + Т-клеток (232). Тем не менее, повышенные уровни липидов, наблюдаемые в нескольких доклинических моделях рака у мышей и у больных раком, оказывают негативное влияние на перекрестное представление ДК. Нормализация уровня липидов в этих моделях рака восстановила функциональную активность богатых липидами ДК и позволила им стать более эффективными при использовании в противораковой вакцине (233). Кроме того, было показано, что анаэробный гликолиз после стимуляции TLR необходим для активации, среди прочего, костимулирующих молекул и IL-12 (234, 235). Также метаболическая характеристика ТАМ коррелирует с их функцией (236, 237). Недавно с использованием генетического подхода была показана прямая связь между метаболизмом ТАМ, сосудистой сетью опухоли и метастазированием. Здесь было показано, что генетическая делеция регулируемого в развитии и ответа на повреждение ДНК 1 (REDD1), ингибитора mTOR, который в высокой степени экспрессируется TAM2, увеличивает поглощение глюкозы и направляет TAM2 на гликолиз (238), отличительный признак TAM1 (239). ). Следовательно, ТАМ конкурировали с опухолевыми эндотелиальными клетками за потребление глюкозы, что приводило к стабилизации сосудистой сети опухоли, тем самым предотвращая метастазирование (238).
Управление различными функциями, стимулирующими развитие опухоли
Несколько ферментов, экспрессируемых TIM, были неразрывно связаны с их иммуносупрессивной функцией, среди которых IDO, ARG-1, iNOS и COX2, и поэтому они были в центре внимания исследований. IDO представляет собой фермент, разлагающий триптофан, который может индуцироваться в MDSC, TADC, TAM и, в некоторой степени, TAN в ответ на стимулы, такие как провоспалительные цитокины, лиганды TLR, гормоны, PGE2 и контактно-зависимые стимулы, такие как B7: цитотоксический T. взаимодействия лимфоцитарного антигена 4 (CTLA-4) (10, 240–242). Чистый эффект опосредованного IDO превращения триптофана в кинуренин заключается в индукции анергии Т-клеток, апоптоза и фиксации CD4 9.0665 + Т-клеток в сторону иммуносупрессивных Treg (243). Эти Treg могут дополнительно усиливать иммуносупрессивную TME, наделяя TADC толерогенными свойствами (например, усиливая IDO), а также рекрутируя и активируя MDSC, которые используют экспрессию IDO, ARG-1 и iNOS в дополнение к иммуносупрессивным цитокинам и ко-ингибирующим молекулы для подавления Т-клеток (244). Таким образом, катаболизм триптофана является важным механизмом ускользания от рака. Экспрессия IDO была связана с плохим клиническим прогнозом при некоторых видах рака, поскольку она коррелирует со снижением выживаемости, а также с повышенным риском метастазирования (245). Следовательно, стратегии нарушения ферментативной активности или экспрессии IDO широко изучались. В исследованиях на животных генетическое ингибирование экспрессии IDO приводило к инфильтрации и активации гранулоцитов, что приводило к TME, способствующему росту опухоли (246). Ингибитор тирозинкиназы иматиниб, который ингибирует активность ЦОГ-2 и, следовательно, блокирует выработку PGE2, что является мощным стимулом для повышения активности IDO, успешно применялся в контексте ингибирования IDO (247, 248). Кроме того, низкомолекулярные ингибиторы IDO проявляют противоопухолевую активность и взаимодействуют с иммунотерапией, лучевой терапией или КТ, вызывая быструю регрессию агрессивных опухолей, в противном случае устойчивых к лечению (249).). Некоторые ингибиторы IDO проходят клиническую оценку в качестве моно- или комбинированной терапии (например, блокада контрольных точек и КТ) (250). Ингибиторами IDO, наиболее интенсивно изучаемыми в клинических испытаниях, являются эпакадостат и индоксимод. В исследовании I фазы эпакадостат нормализовал уровень кинуренина у пациентов с солидными злокачественными новообразованиями на поздних стадиях (251). В исследованиях фазы I-III эпакадостат показал многообещающие результаты в сочетании с различными ингибиторами контрольных точек, включая дурвалумаб (анти-PD-L1), пембролизумаб (анти-PD-1) и ипилимумаб (анти-CTLA-4), при различных видах рака. виды (252). Было показано, что индоксимод противодействует иммуносупрессивному действию кинуренина, активирует множественные иммунные клетки и стимулирует Treg к Т9. 0689 H 1 преобразование (253–255). Текущие клинические исследования фазы II-III изучают влияние индоксимода в комбинации с ингибитором контрольной точки ниволумабом (анти-PD-1) у пациентов с метастатической меланомой. Результатом дальнейшей разработки стало соединение NLG802, пролекарство индоксимода, которое в исследованиях на животных индуцировало противоопухолевый ответ при более низких дозах, чем индоксимод. Недавно NLG802 вошел в фазу I клинических испытаний (256).
ARG-1 — еще один фермент, расщепляющий аминокислоты. Он превращает L-аргинин в L-орнитин и мочевину и может активироваться в MDSC, TAM2 и tolDC под действием множества факторов, включая PGE2 (вырабатываемый COX2), GM-CSF, TGF-β, IL-6 и IL-10. , где последние два действуют через STAT3 (257–260). Также гипоксия связана с экспрессией ARG-1 (261). Экспрессия ARG-1 миелоидными клетками была однозначно связана с активностью, стимулирующей развитие опухоли, и было показано, что L-орнитин дополнительно стимулирует альтернативную активацию миелоидных клеток (262–264). iNOS также является ферментом, превращающим L-аргинин, который в результате производит NO и цитруллин. Экспрессия iNOS была показана в TAM1, воспалительных DC и MDSC в ответ на IL-1β, IL-6, IFN-γ, TNF-α и агонисты TLR4 и была связана как с анти-, так и с проопухолевой активностью (265, 266). Эта двойная роль iNOS и NO, вероятно, зависит от уровней, источника и времени образования NO, а также зависит от TME. На ранних стадиях развития опухоли некоторые миелоидные клетки, такие как макрофаги, могут генерировать высокие концентрации NO, тем самым индуцируя апоптоз опухолевых клеток. Однако при установленном TME миелоидные клетки перепрограммируются, чтобы поддерживать прогрессирование опухоли. В это время низкие количества NO действуют проангиогенно и усиливают рост опухоли и метастазирование, индуцируя факторы роста, такие как VEGF и MMP. Более того, временная связь между экспрессией ARG-1 и iNOS в TIM, таких как макрофаги, была связана с иммуносупрессией. Было высказано предположение, что ARG-1 происходит до экспрессии iNOS, чтобы остановить пролиферацию Т-клеток, и что последующая экспрессия iNOS и высвобождение NO стимулируют апоптоз Т-клеток, тем самым прекращая противораковый иммунный ответ (267). Поэтому были разработаны стратегии блокирования ARG-1 и iNOS. Ингибиторы, такие как нор-N-гидрокси-L-аргинин и аминогуанидин (или 1400W), подавляют активность ARG-1 и iNOS в MDSC, восстанавливают Т-клеточный противоопухолевый иммунитет и замедляют прогрессирование опухоли (176, 268). Более того, недавние исследования показали, что многообещающей стратегией является сочетание ингибирования ARG-1 с терапией иммунных контрольных точек или ЛТ (244, 269).). В нескольких отчетах рекомендуется нацеливаться как на ARG-1, так и на iNOS для усиления терапевтического эффекта (270). Было показано, что ингибиторы фосфодиэстеразы-5 (ФДЭ5), такие как силденафил, тадалафил и варденафил, снижают экспрессию ARG-1 и iNOS. Следовательно, супрессорная активность MDSC была заблокирована, что в конечном итоге усилило опухолеспецифические Т-клеточные ответы (271–273). Точно так же было показано, что высвобождающий NO аспирин снижает ARG-1 и iNOS и увеличивает количество и функцию опухолеспецифических Т-клеток (274). Предшествующее использование ингибиторов ФДЭ-5 и NO-высвобождающего аспирина по другим показаниям побудило их использовать в клинических испытаниях, часто в сочетании с другими методами лечения рака, включая лучевую терапию, КТ и иммунотерапию (вакцинацию, иммуномодулирующие препараты и блокаду иммунных контрольных точек) (275). В то время как многие испытания продолжаются, пилотные испытания у пациентов с метастатической меланомой сообщают о его безопасности и дополнительно показывают снижение частоты и функции MDSC, коррелирующих с повышенной реактивностью Т-клеток и улучшением клинических результатов (276–278). Хотя эти исследования в основном были сосредоточены на роли MDSC, возможно, что эти агенты могут также модулировать другие миелоидные клетки, экспрессирующие ARG-1 и iNOS, такие как TAM и TADC.
COX2 представляет собой фермент, ответственный за метаболическое превращение арахидоновой кислоты в различные простагландины, включая PGE2, который обладает иммуносупрессивной функцией, о чем свидетельствует его способность индуцировать экспрессию IDO и ARG-1 в различных TIM. Другой связью с иммуносупрессивными ферментами, расщепляющими аминокислоты, и ЦОГ2 в миелоидных клетках является наблюдение, что NO участвует в регуляции экспрессии ЦОГ2 (279). Было показано, что ингибирование COX2 в предшественниках миелоидных клеток мыши и человека ингибирует образование MDSC и TAM2 и способствует дифференцировке в направлении зрелых APC (280-283). Генетическая делеция ЦОГ2 в миелоидных клетках 9Было показано, что 0461 in vivo уменьшает инфильтрацию макрофагов, увеличивает число Т-клеток в опухоли и, связанное с этим, замедляет прогрессирование опухоли (284). В различных исследованиях также было показано, что фармакологическое ингибирование ЦОГ2 задерживает рост опухоли и даже синергизирует с иммунотерапией. В этих исследованиях ингибирование ЦОГ-2 было связано со снижением накопления MDSC и снижением экспрессии IDO или ARG-1 (248, 285–288).
В целом, было показано, что ингибирование ключевых иммунодепрессивных ферментов (частично) восстанавливает иммунодепрессивный TME и, таким образом, способствует специфическому иммунитету против рака. Будущие доклинические и клинические исследования должны будут подтвердить, достаточно ли воздействия на один или несколько подавляющих механизмов, вызываемых ТИМ, для контроля прогрессирования рака.
Заключение
В этом обзоре мы подчеркиваем многогранные функции, которыми обладают ТИМ, поддерживающие проявления, прогрессирование и метастазирование рака наряду с препятствиями, которые они образуют на пути к успешным терапевтическим вмешательствам. Об этом свидетельствует обилие публикаций, посвященных ТАМ, TADC, TAN, MDSC и их общим чертам, таким как пластичность и иммуносупрессивные функции. До сих пор были вложены огромные усилия в разработку фармакологических соединений, нацеленных на TIM, чтобы изменить количество их клеток или реполяризовать их функции. Эти стратегии привели к значительным терапевтическим преимуществам в нескольких доклинических моделях и в настоящее время проходят клиническую оценку в качестве дополнительных методов лечения рака.
Тем не менее, требуется осторожность при нейтрализации иммунных факторов, которые не являются уникальными для опухоли и ее TME (например, PGE2, ROS, iNOS или TGF-β), поскольку они необходимы для поддержания гомеостаза. Следовательно, терапия, направленная на истощение TIM или изменение функции, направленная на один из этих факторов, должна быть преходящей или локальной по своей природе. Для будущих методов лечения будет важно расширить наши знания о механизмах, которые приводят в движение иммуносупрессивные баррикады, возводимые ТИМами. Кроме того, следует учитывать тип рака, поскольку некоторые миелоидные композиции могут быть полезными для одного типа и неблагоприятными для другого. Поскольку паттерны миграции и метаболические аберрации возникают на нескольких стадиях прогрессии опухоли, важно подтолкнуть существующую методологию к сочетанию функциональной визуализации и расширенного анализа in silico. Это будет важно для понимания и прогнозирования динамики TME, а также для того, чтобы понять, почему некоторые предсказанные методы лечения неэффективны.0461 in vivo .
Обычные методы лечения часто фокусируются на устранении определенного подмножества среди множества миелоидных клеток, игнорируя те, которые желательны. Лизис опухоли, клиренс и представление антигена являются важными модальностями, которые особенно предназначены для миелоидных клеток, однако они все еще недооценены. Вместо искоренения полной популяции ТИМ мы скорее предлагаем формировать состав миелоидных клеток и состояние поляризации, комбинируя несколько миелоидных иммунотерапий, основываясь на избыточности и общности фенотипа и функции ТИМ. В заключение мы считаем, что реорганизация миелоидного ландшафта может устранить препятствие на пути к успешной терапии рака.
Вклад авторов
Все перечисленные авторы внесли существенный, непосредственный и интеллектуальный вклад в работу и одобрили ее для публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Лаборатория молекулярной и клеточной терапии поддерживается Научным фондом Willy Gepts Университетской клиники Брюсселя; Программа стратегических исследований Свободного университета Брюсселя; Национальный план по борьбе с раком Федерального министерства здравоохранения, Stichting tegen Kanker, Kom op tegen Kanker (Противодействие раку), Фламандское онкологическое общество, Институт науки и инноваций (IWT), Исследовательский фонд Фландрии (FWO-V ), программу финансирования исследований и инноваций Европейского Союза FP7, программу финансирования ERA-NET TRANSCAN и Альянс исследований меланомы. YDV получил грант Эммануэля ван дер Шурена от Kom op tegen Kanker и докторский грант на стратегические фундаментальные исследования от FWO-V. CG является членом Kom op tegen Kanker (Противодействие раку), фламандского онкологического общества.
Ссылки
1. Durgeau A, Virk Y, Corgnac S, Mami-Chouaib F. Недавние достижения в нацеливании CD8 T-клеточного иммунитета для более эффективной иммунотерапии рака. Фронт Иммунол . (2018) 9:14. doi: 10.3389/fimmu.2018.00014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2. Binnewies M, Roberts EW, Kersten K, Chan V, Fearon DF, Merad M, et al. Понимание иммунного микроокружения опухоли (ВРЕМЯ) для эффективной терапии. Нац. мед. (2018) 24:541–50. дои: 10.1038/s41591-018-0014-x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
3. De Vlaeminck Y, Gonzalez-Rascon A, Goyvaerts C, Breckpot K. Миелоидные регуляторные клетки, ассоциированные с раком. Фронт Иммунол. (2016) 7:113. doi: 10.3389/fimmu.2016.00113
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
4. Мантовани А., Аллавена П., Сика А., Балквилл Ф. Воспаление, связанное с раком. Природа (2008) 454:436–44. doi: 10.1038/nature07205
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
5. Ханахан Д., Вайнберг Роберт А. Признаки рака: следующее поколение. Cell (2011) 144:646–74. doi: 10.1016/j.cell.2011.02.013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. Dunn GP, Old LJ, Schreiber RD. Три «Э» иммуноредактирования при раке. Annu Rev Immunol. (2004) 22:329–60. doi: 10.1146/annurev.immunol.22.012703.104803
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
7. Франклин Р.А., Ляо В., Саркар А., Ким М.В., Бивона М.Р., Лю К. и соавт. Клеточное и молекулярное происхождение опухолеассоциированных макрофагов. Наука (2014) 344:921–5. doi: 10.1126/science.1252510
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
8. Тимошук П., Эвенс Х., Марзола В., Вахович К., Васмер М.Х., Датта С. и др. Пролиферация in situ способствует накоплению ассоциированных с опухолью макрофагов в спонтанных опухолях молочной железы. Евро J Иммунол . (2014) 44:2247–62. doi: 10.1002/eji.201344304
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
9. Campbell MJ, Tonlaar NY, Garwood ER, Huo D, Moore DH, Khramtsov AI, et al. Пролиферирующие макрофаги связаны с высокодифференцированным раком молочной железы с отрицательным гормональным рецептором и плохим клиническим исходом. Лечение рака молочной железы . (2011) 128:703–11. doi: 10.1007/s10549-010-1154-y
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
10. Веглиа Ф., Перего М., Габрилович Д. Супрессорные клетки миелоидного происхождения, достигшие совершеннолетия. Нат Иммунол . (2018) 19:108–19. doi: 10.1038/s41590-017-0022-x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11. Zhou J, Nefedova Y, Lei A, Gabrilovich D. Нейтрофилы и PMN-MDSC: их биологическая роль и взаимодействие со стромальными клетками. Семин Иммунол . (2018) 35:19–28. doi: 10.1016/j.smim.2017.12.004
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
12. Лихтенштейн Т., Перес-Янисес Н., Гато М., Калиендо Ф., Кочан Г., Бланко-Лукин И. и др. Высокоэффективная система дифференцировки MDSC, проникающих в опухоль, для обнаружения противоопухолевых мишеней, которая позволяет избежать необходимости создания опухоли у мышей. Oncotarget (2014) 5:7843–57. doi: 10.18632/oncotarget.2279
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
13. Кумар В., Ченг П., Кондамин Т., Мони С., Лангуино Л.Р., Маккефри Дж.С. и др. Фосфатаза CD45 ингибирует активность фактора транскрипции STAT3 в миелоидных клетках и способствует дифференцировке макрофагов, ассоциированных с опухолью. Иммунитет (2016) 44:303–15. doi: 10.1016/j.immuni.2016.01.014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
14. Юн Дж.И., Кумар В., Коллазо М., Нефедова Ю., Кондамин Т., Ченг П. и др. Эпигенетическое молчание гена ретинобластомы регулирует патологическую дифференцировку миелоидных клеток при раке. Нат Иммунол. (2013) 14:211–20. doi: 10.1038/ni.2526
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
15. Сингел К.Л., Сегал Б.Х. Нейтрофилы в микроокружении опухоли: пытаются залечить незаживающую рану. Immunol Rev. (2016) 273:329–43. doi: 10.1111/imr.12459
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16. Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L, et al. Поляризация фенотипа ассоциированных с опухолью нейтрофилов по TGF-бета: «N1» против «N2» TAN. Раковая клетка (2009) 16:183–94. doi: 10.1016/j.ccr.2009.06.017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
17. Кескинов А.А., Шурин М.Р. Миелоидные регуляторные клетки при распространении и метастазировании опухоли. Иммунобиология (2015) 220:236–42. doi: 10.1016/j.imbio.2014.07.017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
18. Coffelt SB, Wellenstein MD, de Visser KE. Нейтрофилы при раке: нейтральные не более. Nat Rev Cancer (2016) 16:431–46. doi: 10.1038/nrc.2016.52
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
19. Дене Н., Мора Дж., Намгаладзе Д., Вейгерт А., Брюн Б. Взаимодействия раковых клеток и макрофагов в микроокружении опухоли. Curr Opin Pharmacol . (2017) 35:12–9. doi: 10.1016/j.coph.2017.04.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
20. Миллс К.Д., Ширер Дж., Эванс Р., Колдуэлл М.Д. Метаболизм аргинина макрофагов и ингибирование или стимуляция рака. Дж Иммунол. (1992) 149:2709–14.
Google Scholar
21. Компакт-диск Миллса. Анатомия открытия: макрофаги M1 и M2. Фронт Иммунол . (2015) 6:212. doi: 10.3389/fimmu.2015.00212
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
22. Van den Bossche J, Bogaert P, van Hengel J, Guerin CJ, Berx G, Movahedi K, et al. Альтернативно активированные макрофаги участвуют в гомотипических и гетеротипических взаимодействиях через IL-4 и индуцированные полиамином комплексы E-кадгерин/катенин. Кровь (2009) 114:4664–74. doi: 10.1182/blood-2009-05-221598
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
23. Ghassabeh GH, De Baetselier P, Brys L, Noel W, Van Ginderachter JA, Meerschaut S, et al. Идентификация общей генной сигнатуры для миелоидных клеток, ассоциированных с цитокинами II типа, выявила in vivo при различных патологических состояниях. Кровь (2006) 108:575–83. doi: 10.1182/blood-2005-04-1485
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
24. Арас С., Заиди М.Р. Предатели без приручения: макрофаги при прогрессировании рака и метастазировании. BrJ Рак (2017) 117:1583–91. doi: 10.1038/bjc.2017.356
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
25. Jinushi M, Chiba S, Yoshiyama H, Masutomi K, Kinoshita I, Dosaka-Akita H, et al. Ассоциированные с опухолью макрофаги регулируют онкогенность и реакцию раковых стволовых/инициирующих клеток на противораковые препараты. Proc Natl Acad Sci USA. (2011) 108:12425–30. doi: 10.1073/pnas.1106645108
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
26. Джеймисон А.М., Дифенбах А., МакМахон К.В., Сюн Н., Карлайл Дж.Р., Раулет Д.Х. Роль иммунорецептора NKG2D в активации иммунных клеток и естественной гибели. Иммунитет (2002) 17:19–29. doi: 10.1016/S1074-7613(02)00333-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
27. Дифенбах А., Джеймисон А.М., Лю С.Д. , Шастри Н., Раулет Д.Х. Лиганды для мышиного рецептора NKG2D: экспрессия опухолевыми клетками и активация NK-клеток и макрофагов. Нат Иммунол. (2000) 1:119–26. doi: 10.1038/77793
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
28. Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL, et al. Активация NK-клеток и Т-клеток с помощью NKG2D, рецептора стресс-индуцируемой MICA. Наука (1999) 285:727–9. doi: 10.1126/science.285.5428.727
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
29. Яннелло А., Томпсон Т.В., Ардолино М., Лоу С.В., Раулет Д.Х. p53-зависимая продукция хемокинов стареющими опухолевыми клетками поддерживает NKG2D-зависимую элиминацию опухоли естественными клетками-киллерами. J Exp Med . (2013) 210:2057–69. doi: 10.1084/jem.20130783
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Kruse PH, Matta J, Ugolini S, Vivier E. Природные рецепторы цитотоксичности и их лиганды. Иммунол Селл Биол . (2014) 92:221–9. doi: 10.1038/icb.2013.98
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
31. Bryceson YT, March ME, Barber DF, Ljunggren HG, Long EO. Поляризация и дегрануляция цитолитических гранул, контролируемая различными рецепторами в покоящихся NK-клетках. J Exp Med. (2005) 202:1001–12. doi: 10.1084/jem.20051143
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
32. Гарридо Ф., Альгарра И. Антигены MHC и опухоль ускользают от иммунного надзора. Adv Cancer Res. (2001) 83:117–58. doi: 10.1016/S0065-230X(01)83005-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
33. Tu MM, Mahmoud AB, Wight A, Mottashed A, Belanger S, Rahim MM, et al. Рецепторы семейства Ly49 необходимы для иммунного надзора за раком, опосредованного естественными клетками-киллерами. Рак Res. (2014) 74:3684–94. doi: 10.1158/0008-5472.CAN-13-3021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
34. Fernandez NC, Lozier A, Flament C, Ricciardi-Castagnoli P, Bellet D, Suter M, et al. Дендритные клетки непосредственно запускают функции NK-клеток: перекрестные помехи связаны с врожденными противоопухолевыми иммунными реакциями in vivo . Нац. мед. (1999) 5:405–11. doi: 10.1038/7403
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
35. Тиба С., Икусима Х., Уэки Х., Янай Х., Кимура Ю., Хангай С. и др. Распознавание опухолевых клеток с помощью Dectin-1 управляет врожденными иммунными клетками для противоопухолевого ответа. Элиф (2014) 3:e04177. doi: 10.7554/eLife.04177
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
36. Ebihara T, Azuma M, Oshiumi H, Kasamatsu J, Iwabuchi K, Matsumoto K, et al. Идентификация полиI:C-индуцируемого мембранного белка, который участвует в опосредованной дендритными клетками активации естественных клеток-киллеров. J Exp Med . (2010) 207:2675–87. doi: 10. 1084/jem.200
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
37. Coulie PG, Van den Eynde BJ, van der Bruggen P, Boon T. Опухолевые антигены, распознаваемые Т-лимфоцитами: в основе иммунотерапии рака. Nat Rev Cancer (2014) 14:135–46. doi: 10.1038/nrc3670
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
38. Маркевич М.А., Караяннопулос Л.Н., Найденко О.В., Мацуи К., Бурак В.Р., Уайз Е.Л., и соавт. Костимуляция с помощью NKG2D усиливает функцию мышиных CD8+ CTL: сходства и различия между костимуляцией NKG2D и CD28. J Иммунол. (2005) 175:2825–33. doi: 10.4049/jimmunol.175.5.2825
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
39. Финн О.Дж. Иммуноонкология: понимание функции и дисфункции иммунной системы при раке. Энн Онкол . (2012) 23 (Приложение 8): viii6–9. doi: 10.1093/annonc/mds256
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
40. Gardai SJ, McPhillips KA, Frasch SC, Janssen WJ, Starefeldt A, Murphy-Ullrich JE, et al. Кальретикулин клеточной поверхности инициирует клиренс жизнеспособных или апоптотических клеток посредством транс-активации LRP на фагоците. Cell (2005) 123:321–34. doi: 10.1016/j.cell.2005.08.032
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
41. Diamond MS, Kinder M, Matsushita H, Mashayekhi M, Dunn GP, Archambault JM, et al. Интерферон типа I избирательно требуется дендритным клеткам для иммунного отторжения опухолей. J Exp Med. (2011) 208:1989–2003. doi: 10.1084/jem.20101158
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
42. Shen YJ, Le Bert N, Chitre AA, Koo CX, Nga XH, Ho SS, et al. Геномная цитозольная ДНК опосредует интерферон-зависимое отторжение I типа клеток В-клеточной лимфомы. Cell Rep. (2015) 11:460–73. doi: 10.1016/j.celrep.2015.03.041
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
43. Chao MP, Jaiswal S, Weissman-Tsukamoto R, Alizadeh AA, Gentles AJ, Volkmer J, et al. Кальретикулин является доминирующим профагоцитарным сигналом при множественных раковых заболеваниях человека и уравновешивается CD47. Sci Transl Med. (2010) 2:63ra94. doi: 10.1126/scitranslmed.3001375
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
44. Вичетти Мигель Р.Д., Черпес Т.Л., Уотсон Л.Дж., МакКенна К.С. Индукция CTL тумороцидной продукции оксида азота внутриопухолевыми макрофагами имеет решающее значение для элиминации опухоли. Дж Иммунол. (2010) 185:6706–18. doi: 10.4049/jimmunol.0
1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
45. Corthay A, Skovseth DK, Lundin KU, Rosjo E, Omholt H, Hofgaard PO, et al. Первичный противоопухолевый иммунный ответ, опосредованный CD4+ Т-клетками. Иммунитет (2005) 22:371–83. doi: 10.1016/j.immuni.2005.02.003
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
46. Matsushita H, Vesely MD, Koboldt DC, Rickert CG, Uppaluri R, Magrini VJ, et al. Анализ экзома рака выявляет Т-клеточно-зависимый механизм иммуноредактирования рака. Природа (2012) 482:400–4. doi: 10.1038/nature10755
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
47. Фон Бемер Л., Маттл М., Боде П., Ландшаммер А., Шафер С., Нубер Н. и др. NY-ESO-1-специфическое иммунологическое давление и побег у пациента с метастатической меланомой. Иммунитет против рака . (2013) 13:12. doi: 10.5167/uzh-79657
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
48. Николау Т., Чен В., Дэвис И.Д., Джексон Х.М., Димопулос Н., Барроу С. и др. Иммунокоррекция и сохранение антигенспецифического иммунитета у пациентов, ранее вакцинированных белком NY-ESO-1, приготовленным в ISCOMATRIX. Рак Иммунол Иммунотер . (2011) 60:1625–37. doi: 10.1007/s00262-011-1041-3
CrossRef Full Text | Академия Google
49. Tchou J, Zhao Y, Levine BL, Zhang PJ, Davis MM, Melenhorst JJ, et al. Безопасность и эффективность внутриопухолевых инъекций Т-клеток химерного антигенного рецептора (CAR) при метастатическом раке молочной железы. Рак Иммунол Рез. (2017) 5:1152–61. doi: 10.1158/2326-6066.CIR-17-0189
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
50. O’Sullivan T, Saddawi-Konefka R, Vermi W, Koebel CM, Arthur C, White JM, et al. Иммуноредактирование рака врожденной иммунной системой при отсутствии адаптивного иммунитета. J Exp Med. (2012) 209:1869–82. doi: 10.1084/jem.20112738
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
51. Арендс Т., Борст Дж. Противоположные роли CD4(+) Т-клеток в противоопухолевом иммунитете. Immunology (2018) 154. doi: 10.1111/imm.12941
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
52. Бен-Меир К., Твайк Н., Банияш М. Пластичность и биологическое разнообразие миелоидных супрессорных клеток. Карр Опин Иммунол. (2018) 51:154–61. doi: 10.1016/j.coi.2018.03.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
53. Миттал Д., Губин М.М., Шрайбер Р.Д., Смит М.Дж. Новый взгляд на иммуноредактирование рака и его три фазы: элиминацию, равновесие и побег. Карр Опин Иммунол. (2014) 27:16–25. doi: 10.1016/j.coi.2014.01.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
54. Крыско Д.В., Гарг А.Д., Качмарек А., Крыско О., Агостинис П., Ванденабиле П. Иммуногенная гибель клеток и DAMP в терапии рака. Nat Rev Cancer (2012) 12:860–75. doi: 10.1038/nrc3380
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
55. Braumuller H, Wieder T, Brenner E, Assmann S, Hahn M, Alkhaled M, et al. Цитокины Т-хелперов-1 вызывают старение рака. Природа (2013) 494:361–5. doi: 10.1038/nature11824
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
56. Курсунель М.А., Эсендальи Г. Нерассказанная история IFN-gamma в биологии рака. Фактор роста цитокинов Ред. (2016) 31:73–81. doi: 10.1016/j.cytogfr.2016.07.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
57. Yang H, Lundback P, Ottosson L, Erlandsson-Harris H, Venereau E, Bianchi ME, et al. Окислительно-восстановительная модификация остатков цистеина регулирует активность цитокинов высокоподвижной группы box-1 (HMGB1). Мол мед. (2012) 18:250–9. doi: 10.2119/molmed.2011.00389
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
58. Эль-Кенави А., Раффелл Б. Воспаление, АФК и мутагенез. Раковая клетка . (2017) 32:727–9. doi: 10.1016/j.ccell.2017.11.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
59. Roca H, Jones JD, Purica MC, Weidner S, Koh AJ, Kuo R, et al. Индуцированный апоптозом CXCL5 ускоряет воспаление и рост метастазов опухоли предстательной железы в кости. Дж Клин Инвест . (2017) 128: 248–66. doi: 10.1172/JCI
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
60. Tamadaho RSE, Hoerauf A, Layland LE. Иммуномодулирующие эффекты супрессорных клеток миелоидного происхождения при заболеваниях: роль в развитии рака и инфекций. Иммунобиология (2018) 223:432–42. doi: 10.1016/j.imbio.2017.07.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
61. Мохамед Э., Аль-Хами А.А., Родригес П.С. Клеточный метаболический ландшафт в опухолевой среде регулирует активность миелоидных инфильтратов. Селл Мол Иммунол . (2018) 15:421–7. doi: 10.1038/s41423-018-0001-7
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
62. Шрайбер Р.Д., Олд Л.Дж., Смит М.Дж. Иммуноредактирование рака: интеграция роли иммунитета в подавлении и развитии рака. Наука (2011) 331:1565–70. doi: 10.1126/science.1203486
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
63. Лин Э.Ю., Гуон-Эванс В., Нгуен А.В., Поллард Дж.В. Фактор роста макрофагов CSF-1 в развитии молочной железы и опухолевой прогрессии. J Биол Неоплазия молочной железы (2002) 7:147–62. doi: 10.1023/A:1020399802795
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
64. Zhang X, Xue D, Hao F, Xie L, He J, Gai J и др. Сверхэкспрессия фактора ремоделирования и спейсинга 1 связана с плохим прогнозом при почечно-клеточной карциноме. Онкол Летт . (2018) 15:3852–7. doi: 10.3892/ol.2018.7797
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
65. Чималь-Рамирес Г.К., Эспиноза-Санчес Н.А., Фуэнтес-Панана Э.М. Проопухолевая активность иммунного ответа: понимание механизмов иммунологического сдвига, онкотренинга и онкопромотирования. Дж Онкол . (2013) 2013: 835956. doi: 10.1155/2013/835956
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
66. Dufait I, Van Valckenborgh E, Menu E, Escors D, De Ridder M, Breckpot K. Преобразователь сигнала и активатор транскрипции 3 в супрессорных клетках миелоидного происхождения: возможность для лечения рака. Oncotarget (2016) 7:42698–715. doi: 10.18632/oncotarget.8311
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
67. Кортылевский М., Ю. Х. Роль Stat3 в подавлении противоопухолевого иммунитета. Курр Опин Иммунол. (2008) 20:228–33. doi: 10.1016/j.coi.2008.03.010
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
68. Arce F, Breckpot K, Stephenson H, Karwacz K, Ehrenstein MR, Collins M, et al. Селективная активация ERK дифференцирует толерогенные дендритные клетки мыши и человека, увеличивает количество антиген-специфических регуляторных Т-клеток и подавляет экспериментальный воспалительный артрит. Ревматоидный артрит. (2011) 63:84–95. doi: 10.1002/art.30099
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
69. Maenhout SK, Van Lint S, Emeagi PU, Thielemans K, Aerts JL. Повышенная супрессивная способность инфильтрирующих опухоль супрессорных клеток миелоидного происхождения по сравнению с их периферическими аналогами. Int J Cancer (2014) 134:1077–90. doi: 10.1002/ijc.28449
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
70. Ballbach M, Dannert A, Singh A, Siegmund DM, Handgretinger R, Piali L, et al. Экспрессия молекул контрольной точки на супрессорных клетках миелоидного происхождения. Иммунол Летт . (2017) 192:1–6. doi: 10.1016/j.imlet.2017.10.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
71. Прима В., Калиберова Л.Н., Калиберов С., Куриел Д.Т., Кусмарцев С. Путь COX2/mPGES1/PGE2 регулирует экспрессию PD-L1 в опухолеассоциированных макрофагах и супрессорных клетках миелоидного происхождения. Proc Natl Acad Sci USA. (2017) 114:1117–22. doi: 10.1073/pnas.16124
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
72. Такахаси Х., Сакакура К., Кудо Т., Тойода М., Кайра К., Ояма Т. и др. Связанные с раком фибробласты способствуют созданию иммуносупрессивной микросреды за счет индукции и накопления проопухолевых макрофагов. Oncotarget (2017) 8:8633–47. doi: 10.18632/oncotarget.14374
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
73. Chen J, Ye Y, Liu P, Yu W, Wei F, Li H, et al. Подавление Т-клеток супрессорными клетками миелоидного происхождения при раке. Хум Иммунол. (2017) 78:113–9. doi: 10.1016/j.humimm.2016.12.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
74. Mougiakakos D, Jitschin R, von Bahr L, Poschke I, Gary R, Sundberg B, et al. Иммуносупрессивные CD14+HLA-DRlow/neg IDO+ миелоидные клетки у пациентов после аллогенной трансплантации гемопоэтических стволовых клеток. Лейкемия (2013) 27:377–88. doi: 10.1038/leu.2012.215
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
75. Yu J, Du W, Yan F, Wang Y, Li H, Cao S, et al. Супрессорные клетки миелоидного происхождения подавляют противоопухолевый иммунный ответ за счет экспрессии IDO и коррелируют с метастазированием в лимфатические узлы у пациентов с раком молочной железы. J Иммунол. (2013) 190:3783–97. doi: 10.4049/jimmunol.13
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
153. Nywening TM, Belt BA, Cullinan DR, Panni RZ, Han BJ, Sanford DE, et al. Воздействие как на связанные с опухолью CXCR2 + нейтрофилы, так и на CCR2 + макрофаги нарушает рекрутирование миелоидных клеток и улучшает химиотерапевтические реакции при аденокарциноме протоков поджелудочной железы. Гут (2017) 67:1112–3. doi: 10.1136/gutjnl-2017-313738
Полный текст CrossRef | Google Scholar
154.
Ruffell B, Coussens LM. Макрофаги и терапевтическая резистентность при раке. Раковая клетка. (2015) 27:462–72. doi: 10.1016/j.ccell.2015.02.015PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
155. Хорикава Н., Абико К., Мацумура Н., Хаманиши Дж., Баба Т., Ямагути К. и др. Экспрессия сосудистого эндотелиального фактора роста при раке яичников ингибирует опухолевый иммунитет за счет накопления супрессорных клеток миелоидного происхождения. Clin Cancer Res. (2017) 23:587–99. doi: 10.1158/1078-0432.CCR-16-0387
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
156. Linde N, Lederle W, Depner S, van Rooijen N, Gutschalk CM, Mueller MM. Канцерогенез кожи, индуцированный фактором роста эндотелия сосудов, зависит от рекрутирования и альтернативной активации макрофагов. Дж. Патол. (2012) 227:17–28. doi: 10.1002/path.3989
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
157.
Avraham-Davidi I, Yona S, Grunewald M, Landsman L, Cochain C, Silvestre JS, et al. Локальное образование рекрутированных VEGF моноцитов улучшает их работу в качестве ангиогенных и артериогенных вспомогательных клеток. J Exp Med. (2013) 210:2611–25. doi: 10.1084/jem.20120690PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
158. Chen XW, Sun JG, Zhang LP, Liao XY, Liao RX. Рекрутирование моноцитов CD11b(+)Ly6C(+) в ксенотрансплантатах немелкоклеточного рака легкого, зараженных антителом против VEGF. Онкол Летт . (2017) 14: 615–22. doi: 10.3892/ol.2017.6236
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
159. Д’Агостино Р.Б. Старший. Изменение конечных точек при одобрении препарата для лечения рака груди – история Avastin. N Engl J Med. (2011) 365:e2. doi: 10.1056/NEJMp1106984
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
160. Gilbert MR, Dignam JJ, Armstrong TS, Wefel JS, Blumenthal DT, Vogelbaum MA, et al.
Рандомизированное исследование бевацизумаба при недавно диагностированной глиобластоме. N Engl J Med. (2014) 370:699–708. doi: 10.1056/NEJMoa1308573PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
161. Du Four S, Maenhout SK, Benteyn D, De Keersmaecker B, Duerinck J, Thielemans K, et al. Прогрессирование заболевания у пациентов с рецидивирующей глиобластомой, получавших лечение акситинибом, ингибитором VEGFR, связано с увеличением числа регуляторных Т-клеток и истощением Т-клеток. Рак Иммунол Иммунотер . (2016) 65:727–40. doi: 10.1007/s00262-016-1836-3
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
162. Heine A, Held SA, Daecke SN, Riethausen K, Kotthoff P, Flores C, et al. Ингибитор VEGF-рецепторов акситиниб нарушает фенотип и функцию дендритных клеток. PLoS ONE (2015) 10:e0128897. doi: 10.1371/journal.pone.0128897
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
163.
Du Four S, Maenhout SK, De Pierre K, Renmans D, Niclou SP, Thielemans K, et al. Акситиниб увеличивает инфильтрацию иммунных клеток и снижает супрессивную способность моноцитарных MDSC в модели внутричерепной меланомы у мышей. Онкоиммунология (2015) 4:e998107. doi: 10.1080/2162402X.2014.998107PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
164. Nishioka Y, Hirao M, Robbins PD, Lotze MT, Tahara H. Индукция системного и терапевтического противоопухолевого иммунитета с помощью внутриопухолевой инъекции дендритных клеток, генетически модифицированных для экспрессии интерлейкина 12. Cancer Res. (1999) 59:4035–41.
Реферат PubMed | Google Scholar
165. Mazzolini G, Alfaro C, Sangro B, Feijoo E, Ruiz J, Benito A, et al. Внутриопухолевая инъекция дендритных клеток, сконструированных для секреции интерлейкина-12 с помощью рекомбинантного аденовируса, у пациентов с метастатическими карциномами желудочно-кишечного тракта. J Клин Онкол.
(2005) 23:999–1010. doi: 10.1200/JCO.2005.00.463PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
166. Стируолт Д.Л., Радич Дж.П. Роль FLT3 в злокачественных новообразованиях кроветворной системы. Nat Rev Рак . (2003) 3:650–65. doi: 10.1038/nrc1169
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
167. Ватович С.С., Лю Ю.Дж. Механизмы, регулирующие спецификацию и развитие дендритных клеток. Иммунол Ред. (2010) 238:76–92. doi: 10.1111/j.1600-065X.2010.00949.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
168. Линч Д.Х., Андреасен А., Марасковский Э., Уитмор Дж., Миллер Р.Э., Шух Дж.К. Лиганд Flt3 индуцирует регрессию опухоли и противоопухолевый иммунный ответ in vivo . Нац. мед. (1997) 3:625–31. doi: 10.1038/nm0697-625
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
169. Эше С., Субботин В.М., Малишевский С.
, Лотце М.Т., Шурин М.Р. Введение лиганда FLT3 ингибирует рост опухоли при мышиной меланоме и лимфоме. Рак Res. (1998) 58:380–3.Реферат PubMed | Google Scholar
170. Hou S, Kou G, Fan X, Wang H, Qian W, Zhang D, et al. Избавление от гепатомы и рака толстой кишки у мышей с помощью генной терапии Flt3L в сочетании с 5-ФУ. Рак Иммунол Иммунотер . (2007) 56:1605–13. doi: 10.1007/s00262-007-0306-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
171. Tandon M, Vemula SV, Sharma A, Ahi YS, Mittal S, Bangari DS, et al. Апоптоз, опосредованный взаимодействием EphrinA1-EphA2, и иммунотерапия, индуцированная Flt3L, ингибируют рост опухоли в мышиной модели рака молочной железы. J Gene Med. (2012) 14:77–89. doi: 10.1002/jgm.1649
CrossRef Полный текст | Google Scholar
172. Fong L, Hou Y, Rivas A, Benike C, Yuen A, Fisher GA, et al. Вакцинация измененным пептидным лигандом с лигандом Flt3 размножала дендритные клетки для иммунотерапии опухолей.
Proc Natl Acad Sci USA. (2001) 98:8809–14. doi: 10.1073/pnas.141226398PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
173. Senzer NN, Kaufman HL, Amatruda T, Nemunaitis M, Reid T, Daniels G, et al. Клинические испытания фазы II онколитического герпесвируса второго поколения, кодирующего гранулоцитарно-макрофагальный колониестимулирующий фактор, у пациентов с нерезектабельной метастатической меланомой. J Клин Онкол . (2009) 27:5763–71. doi: 10.1200/JCO.2009.24.3675
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
174. Kaufman HL, Kim DW, DeRaffele G, Mitcham J, Coffin RS, Kim-Schulze S. Местный и отдаленный иммунитет, индуцированный внутриочаговой вакцинацией онколитическим вирусом герпеса, кодирующим GM-CSF, у пациентов со стадиями IIIc и IV. меланома. Энн Сург Онкол . (2010) 17:718–30. doi: 10.1245/s10434-009-0809-6
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
175.
Лихтенштейн Т., Перес-Янисес Н., Бланко-Лукин И., Гойвартс С., Шварце Дж., Дюфаит И. и соавт. Вакцины против меланомы, сконструированные для одновременной модуляции примирования цитокинов и молчания PD-L1, охарактеризованы с использованием ex vivo супрессорных клеток миелоидного происхождения в качестве показателя терапевтической эффективности. Онкоиммунология (2014) 3:e945378. doi: 10.4161/21624011.2014.945378PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
176. Dufait I, Schwarze JK, Liechtenstein T, Leonard W, Jiang H, Escors D, et al. Ex vivo поколение супрессорных клеток миелоидного происхождения, которые моделируют иммуносупрессивную среду опухоли при колоректальном раке. Онкотаргет. (2015) 6:12369–82. doi: 10.18632/oncotarget.3682
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
177. De Santo C, Salio M, Masri SH, Lee LY-H, Dong T, Speak AO и др. Инвариантные NKT-клетки снижают иммуносупрессивную активность супрессорных клеток миелоидного происхождения, индуцированных вирусом гриппа А, у мышей и людей.
Дж Клин Инвест. (2008) 118:4036–48. doi: 10.1172/JCI36264PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
178. Широта Ю., Широта Х., Клинман Д.М. Внутриопухолевая инъекция олигонуклеотидов CpG индуцирует дифференцировку и снижает иммуносупрессивную активность супрессорных клеток миелоидного происхождения. Дж Иммунол. (2012) 188:1592–9. doi: 10.4049/jimmunol.1101304
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
179. Takeda Y, Kataoka K, Yamagishi J, Ogawa S, Seya T, Matsumoto M. Адъювант, специфичный для TLR3, снимает врожденную устойчивость к блокаде PD-L1 без токсичности цитокинов при иммунотерапии противоопухолевой вакциной. Cell Rep. (2017) 19:1874–87. doi: 10.1016/j.celrep.2017.05.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
180. Shime H, Matsumoto M, Seya T. Двухцепочечная РНК способствует CTL-независимому цитолизу опухоли, опосредованному внутриопухолевыми миелоидными клетками CD11b(+)Ly6G(+) через сигнальный путь TICAM-1.
Гибель клеток Отличие . (2017) 24:385–96. doi: 10.1038/cdd.2016.131PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
181. Zoglmeier C, Bauer H, Norenberg D, Wedekind G, Bittner P, Sandholzer N, et al. CpG блокирует иммуносупрессию супрессорными клетками миелоидного происхождения у мышей с опухолями. Clin Cancer Res. (2011) 17:1765–75. doi: 10.1158/1078-0432.CCR-10-2672
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
182. Shime H, Matsumoto M, Oshiumi H, Tanaka S, Nakane A, Iwakura Y, et al. Передача сигналов Toll-подобного рецептора 3 превращает миелоидные клетки, поддерживающие опухоль, в онкоцидные эффекторы. Proc Natl Acad Sci USA . (2012) 109:2066–71. doi: 10.1073/pnas.1113099109
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
183. Matsumoto M, Tatematsu M, Nishikawa F, Azuma M, Ishii N, Morii-Sakai A, et al. Определенный TLR3-специфический адъювант, который индуцирует активацию NK и CTL без значительной продукции цитокинов in vivo .
Нац.коммун. (2015) 6:6280. doi: 10.1038/ncomms7280PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
184. Bunt SK, Clements VK, Hanson EM, Sinha P, Ostrand-Rosenberg S. Воспаление усиливает перекрестные помехи миелоидных супрессорных клеток путем передачи сигналов через Toll-подобный рецептор 4. J Leukoc Biol. (2009) 85:996–1004. doi: 10.1189/jlb.0708446
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
185. Ko HJ, Lee JM, Kim YJ, Kim YS, Lee KA, Kang CY. Иммуносупрессивные супрессорные клетки миелоидного происхождения могут быть преобразованы в иммуногенные АПК с помощью активированных NKT-клеток: альтернативной противоопухолевой вакцины на клеточной основе. J Иммунол. (2009) 182:1818–28. doi: 10.4049/jimmunol.0802430
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
186. De Santo C, Arscott R, Booth S, Karydis I, Jones M, Asher R, et al. Инвариантные NKT-клетки модулируют супрессивную активность нейтрофилов, секретирующих IL-10, дифференцированных по амилоиду А сыворотки.
Нат Иммунол . (2010) 11:1039–46. doi: 10.1038/ni.1942CrossRef Полный текст | Google Scholar
187. Beatty GL, Chiorean EG, Fishman MP, Saboury B, Teitelbaum UR, Sun W, et al. Агонисты CD40 изменяют строму опухоли и проявляют эффективность против карциномы поджелудочной железы у мышей и людей. Наука (2011) 331:1612–6. doi: 10.1126/science.1198443
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
188. Beatty GL, Torigian DA, Chiorean EG, Saboury B, Brothers A, Alavi A, et al. Фаза I исследования моноклонального антитела-агониста CD40 (CP-870,893) в комбинации с гемцитабином у пациентов с распространенной аденокарциномой протоков поджелудочной железы. Клин Рак Res . (2013) 19:6286–95. doi: 10.1158/1078-0432.CCR-13-1320
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
189. Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Лигирование CD40 на дендритных клетках вызывает выработку высоких уровней интерлейкина-12 и усиливает стимулирующую способность Т-клеток: помощь Т-Т через APC активация.
J Exp Med. (1996) 184:747–52. doi: 10.1084/jem.184.2.747PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
190. Scarlett UK, Cubillos-Ruiz JR, Nesbeth YC, Martinez DG, Engle X, Gewirtz AT, et al. На месте стимуляция CD40 и Toll-подобного рецептора 3 трансформирует инфильтрирующие рак яичников дендритные клетки из иммуносупрессивных в иммуностимулирующие клетки. Рак Res. (2009) 69:7329–37. doi: 10.1158/0008-5472.CAN-09-0835
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
191. Van Lint S, Renmans D, Broos K, Goethals L, Maenhout S, Benteyn D, et al. Внутриопухолевая доставка мРНК TriMix приводит к активации Т-клеток путем перекрестного представления дендритных клеток. Рак Иммунол Рез. (2016) 4:146–56. doi: 10.1158/2326-6066.CIR-15-0163
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
192. Cavallo F, Giovarelli M, Forni G, Signorelli P, Musiani P, Modesti A, et al.
Противоопухолевая эффективность клеток аденокарциномы, сконструированных для продукции интерлейкина 12 (IL-12) или других цитокинов, по сравнению с экзогенным IL-12. JNCI. J Natl Cancer Inst . (1997) 89:1049–58. doi: 10.1093/jnci/89.14.1049PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
193. Tanriover N, Ulu MO, Sanus GZ, Bilir A, Canbeyli R, Oz B, et al. Эффекты системного и внутриопухолевого лечения интерлейкином-12 в модели глиомы крыс С6. Нейрол Рез . (2008) 30:511–7. doi: 10.1179/174313208X289516
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
194. Керкар С.П., Гольдшмид Р.С., Мурански П., Чиннасами Д., Ю З., Регер Р.Н. и соавт. IL-12 запускает программные изменения в дисфункциональных клетках миелоидного происхождения в опухолях мышей. Дж Клин Инвест . (2011) 121:4746–57. doi: 10.1172/JCI58814
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
195.
Керкар С.П., Рестифо Н.П. Мощность и подводные камни Ил-12. Кровь (2012) 119:4096. doi: 10.1182/blood-2012-03-415018PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
196. Уоткинс С.К., Эгильмез Н.К., Саттлз Дж., Стаут Р.Д. IL-12 быстро изменяет функциональный профиль опухолеассоциированных и инфильтрирующих опухоль макрофагов in vitro и in vivo . Дж Иммунол. (2007) 178:1357–62. doi: 10.4049/jimmunol.178.3.1357
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
197. Haicheur N, Escudier B, Dorval T, Negrier S, De Mulder PH, Dupuy JM, et al. Индукция цитокинов и рецепторов растворимых цитокинов после введения IL-12 у онкологических больных. Clin Exp Immunol. (2000) 119:28–37. doi: 10.1046/j.1365-2249.2000.01112.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
198. Portielje JE, Lamers CH, Kruit WH, Sparreboom A, Bolhuis RL, Stoter G, et al.
Повторные введения интерлейкина (ИЛ)-12 связаны с постоянно повышенными уровнями ИЛ-10 в плазме и снижением ответов ИФН-гамма, фактора некроза опухоли-альфа, ИЛ-6 и ИЛ-8. Клин Рак Res . (2003) 9:76–83.Реферат PubMed | Google Scholar
199. Yin XL, Wang N, Wei X, Xie GF, Li JJ, Liang HJ. Интерлейкин-12 ингибирует выживание стволовых клеток рака толстой кишки человека in vitro и их способность инициировать опухоль у мышей. Рак Летт . (2012) 322:92–7. doi: 10.1016/j.canlet.2012.02.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
200. Gollob JA, Mier JW, Veenstra K, McDermott DF, Clancy D, Clancy M, et al. Испытание фазы I внутривенного введения интерлейкина-12 два раза в неделю у пациентов с метастатическим почечно-клеточным раком или злокачественной меланомой: способность поддерживать индукцию IFN-гамма связана с клиническим ответом. Клин Рак Рез . (2000) 6:1678–92.
Реферат PubMed | Google Scholar
201.
Motzer RJ, Rakhit A, Schwartz LH, Olencki T, Malone TM, Sandstrom K, et al. Испытание фазы I подкожного рекомбинантного человеческого интерлейкина-12 у пациентов с распространенным почечно-клеточным раком. Клин Рак Res . (1998) 4:1183–91.Реферат PubMed | Google Scholar
202. Cebon J, Jager E, Shackleton MJ, Gibbs P, Davis ID, Hopkins W, et al. Два исследования фазы I рекомбинантного человеческого IL-12 в низких дозах с меланом-А и пептидами гриппа у субъектов с прогрессирующей злокачественной меланомой. Иммунитет против рака . (2003) 3:7.
Реферат PubMed | Google Scholar
203. Лоренци С., Маттеи Ф., Систигу А., Браччи Л., Спадаро Ф., Санчес М. и др. IFN типа I контролируют удержание антигена и выживание CD8альфа(+) дендритных клеток после поглощения опухолевыми апоптотическими клетками, что приводит к перекрестному праймированию. Дж Иммунол . (2011) 186:5142–50. doi: 10.4049/jimmunol.1004163
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
204.
Fuertes MB, Kacha AK, Kline J, Woo SR, Kranz DM, Murphy KM, et al. Сигналы IFN-хозяина типа I необходимы для противоопухолевого ответа CD8+ Т-клеток через CD8{альфа}+ дендритные клетки. J Exp Med. (2011) 208:2005–16. doi: 10.1084/jem.20101159PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
205. Picozzi VJ, Abrams RA, Decker PA, Traverso W, O’Reilly EM, Greeno E, et al. Многоцентровое исследование фазы II адъювантной терапии резецированного рака поджелудочной железы с использованием цисплатина, 5-фторурацила и химиолучевой терапии на основе интерферона-альфа-2b: исследование ACOSOG Z05031. Энн Онкол . (2011) 22:348–54. doi: 10.1093/annonc/mdq384
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
206. Van der Jeught K, Joe PT, Bialkowski L, Heirman C, Daszkiewicz L, Liechtenstein T, et al. Внутриопухолевое введение мРНК, кодирующей фузокин, состоящий из ИФН-бета и эктодомена рецептора II ТФР-бета, потенцирует противоопухолевый иммунитет.
Oncotarget (2014) 5:10100–13. doi: 10.18632/oncotarget.2463PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
207. Tanikawa T, Wilke CM, Kryczek I, Chen GY, Kao J, Nunez G, et al. Удаление интерлейкина-10 способствует развитию, росту и метастазированию опухоли. Рак Res. (2012) 72:420–9. doi: 10.1158/0008-5472.CAN-10-4627
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
208. Брекпот К., Эскорс Д. Дендритные клетки для активной противораковой иммунотерапии: нацеливание на пути активации посредством генетической модификации. Целевые лекарственные средства Endocr Metab для лечения иммунных расстройств. (2009) 9:328–43. doi: 10.2174/187153009789839156
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
209. Arce F, Kochan G, Breckpot K, Stephenson H, Escors D. Избирательная активация внутриклеточных сигнальных путей в дендритных клетках для иммунотерапии рака.
Противораковые препараты Med Chem . (2012) 12:29–39. doi: 10.2174/187152012798764679PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
210. Maenhout SK, Du Four S, Corthals J, Neyns B, Thielemans K, Aerts JL. AZD1480 задерживает рост опухоли в модели меланомы, одновременно усиливая супрессорную активность супрессорных клеток миелоидного происхождения. Oncotarget (2014) 5:6801–15. doi: 10.18632/oncotarget.2254
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
211. He W, Zhu Y, Mu R, Xu J, Zhang X, Wang C, et al. Jak2-селективный ингибитор сильно меняет иммунную супрессию, модулируя микроокружение опухоли для иммунотерапии рака. Биохим Фармакол . (2017) 145:132–46. doi: 10.1016/j.bcp.2017.08.019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
212. Spinetti T, Spagnuolo L, Mottas I, Secondini C, Treinies M, Rüegg C, et al. Иммунотерапия рака на основе TLR7 уменьшает количество внутриопухолевых супрессорных клеток миелоидного происхождения и блокирует их иммуносупрессивную функцию.
Онкоиммунология (2016) 5:e1230578. doi: 10.1080/2162402X.2016.1230578PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
213. Кортылевски М., Морейра Д. Миелоидные клетки как мишень для олигонуклеотидной терапии: превращение препятствий в возможности. Рак Иммунол Иммунотер . (2017) 66:979–88. doi: 10.1007/s00262-017-1966-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
214. Fleming V, Hu X, Weber R, Nagibin V, Groth C, Altevogt P, et al. Нацеливание на супрессорные клетки миелоидного происхождения для обхода индуцированной опухолью иммуносупрессии. Фронт Иммунол . (2018) 9:398. doi: 10.3389/fimmu.2018.00398
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
215. Luo Z, Wang C, Yi H, Li P, Pan H, Liu L, et al. Нановавакцина, нагруженная поли I:C и STAT3 siRNA, надежно вызывает противоопухолевый иммунный ответ посредством модуляции ассоциированных с опухолью дендритных клеток in vivo .
Биоматериалы (2015) 38:50–60. doi: 10.1016/j.biomaterials.2014.10.050PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
216. Zhang Q, Hossain DM, Duttagupta P, Moreira D, Zhao X, Won H, et al. Устойчивая к сыворотке приманка CpG-STAT3 для нацеливания на выживание и передачу сигналов иммунных контрольных точек при остром миелоидном лейкозе. Кровь (2016) 127:1687–700. doi: 10.1182/blood-2015-08-665604
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
217. Бисвас С.К., Тергаонкар В. Миелоидный фактор дифференцировки, независимый от фактора 88 Toll-подобного рецепторного пути: поддержание воспаления или повышение толерантности? Int J Biochem Cell Biol . (2007) 39:1582–92. doi: 10.1016/j.biocel.2007.04.021
CrossRef Полный текст | Google Scholar
218. Осима Х., Осима М., Инаба К., Такето М.М. Гиперпластические опухоли желудка, индуцированные активированными макрофагами у трансгенных мышей COX-2/mPGES-1.
Эмбо J . (2004) 23:1669–78. doi: 10.1038/sj.emboj.7600170PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
219. Hagemann T, Lawrence T, McNeish I, Charles KA, Kulbe H, Thompson RG, et al. «Переобучение» ассоциированных с опухолью макрофагов путем нацеливания на NF-kappaB. J Exp Med. (2008) 205:1261–8. doi: 10.1084/jem.20080108
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
220. Томпсон М.Г., Ларсон М., Видрин А., Барриос К., Наварро Ф., Мейерс К. и др. Белковые комплексы FOXO3-NF-kappaB RelA снижают передачу сигналов и функцию провоспалительных клеток. Дж Иммунол . (2015) 195:5637–47. doi: 10.4049/jimmunol.1501758
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
221. Breckpot K, Aerts-Toegaert C, Heirman C, Peeters U, Beyaert R, Aerts JL, et al. Ослабленная экспрессия А20 заметно повышает эффективность активированных двухцепочечной РНК дендритных клеток в качестве противораковой вакцины.
J Иммунол. (2009) 182:860–70. doi: 10.4049/jimmunol.182.2.860PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
222. Kaneda MM, Messer KS, Ralainirina N, Li H, Leem CJ, Gorjestani S, et al. PI3Kgamma — это молекулярный переключатель, контролирующий подавление иммунитета. Природа (2016) 539:437–42. doi: 10.1038/nature19834
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
223. De Henau O, Rausch M, Winkler D, Campesato LF, Liu C, Cymerman DH, et al. Преодоление устойчивости к терапии блокадой контрольных точек путем нацеливания на PI3Kgamma в миелоидных клетках. Природа (2016) 539:443–7. doi: 10.1038/nature20554
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
224. Covarrubias AJ, Aksoylar HI, Yu J, Snyder NW, Worth AJ, Iyer SS, et al. Передача сигналов Akt-mTORC1 регулирует Acly, чтобы интегрировать метаболический вход в контроль активации макрофагов. Элайф (2016) 5:e11612.
doi: 10.7554/eLife.11612PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
225. Nicodeme E, Jeffrey KL, Schaefer U, Beinke S, Dewell S, Chung CW, et al. Подавление воспаления синтетическим миметиком гистонов. Природа (2010) 468:1119–23. doi: 10.1038/nature09589
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
226. Wang H, Cheng F, Woan K, Sahakian E, Merino O, Rock-Klotz J, et al. Ингибитор гистондеацетилазы LAQ824 усиливает воспалительные реакции в макрофагах посредством регуляции транскрипции IL-10. Дж Иммунол. (2011) 186:3986–96. doi: 10.4049/jimmunol.1001101
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
227. Chen X, Barozzi I, Termanini A, Prosperini E, Recchiuti A, Dalli J, et al. Потребность в гистондеацетилазе Hdac3 для программы экспрессии воспалительного гена в макрофагах. Proc Natl Acad Sci USA . (2012) 109:E2865–74. doi: 10.1073/pnas.1121131109
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
228.
Guerriero JL, Sotayo A, Ponichtera HE, Castrillon JA, Pourzia AL, Schad S, et al. Ингибирование HDAC класса IIa уменьшает опухоль молочной железы и метастазы за счет противоопухолевых макрофагов. Природа (2017) 543:428–32. doi: 10.1038/nature21409PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
229. Аль-Хами А.А., Родригес П.С., Очоа А.С. Пути энергетического метаболизма контролируют судьбу и функцию миелоидных иммунных клеток. J Лейкок Биол . (2017) 102: 369–80. doi: 10.1189/jlb.1VMR1216-535R
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
230. Al-Khami AA, Zheng L, Del Valle L, Hossain F, Wyczechowska D, Zabaleta J, et al. Экзогенное поглощение липидов индуцирует метаболическое и функциональное перепрограммирование опухолеассоциированных супрессорных клеток миелоидного происхождения. Онкоиммунология (2017) 6:e1344804. doi: 10.1080/2162402X.2017.1344804
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
231.
Hossain F, Al-Khami AA, Wyczechowska D, Hernandez C, Zheng L, Reiss K, et al. Ингибирование окисления жирных кислот модулирует иммуносупрессивные функции супрессорных клеток миелоидного происхождения и усиливает терапию рака. Рак Иммунол Рез . (2015) 3:1236–47. doi: 10.1158/2326-6066.CIR-15-0036PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
232. Bougneres L, Helft J, Tiwari S, Vargas P, Chang BH, Chan L, et al. Роль липидных тел в перекрестной презентации фагоцитированных антигенов МНС класса I в дендритных клетках. Иммунитет (2009) 31:232–44. doi: 10.1016/j.immuni.2009.06.022
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
233. Гербер Д.Л., Цао В., Нефедова Ю., Новицкий С.В., Нагарадж С., Тюрин В.А., и соавт. Накопление липидов и дисфункция дендритных клеток при раке. Нац. мед. (2010) 16:880–6. doi: 10.1038/nm.2172
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
234.
Krawczyk CM, Holowka T, Sun J, Blagih J, Amiel E, DeBerardinis RJ, et al. Индуцированные толл-подобными рецепторами изменения гликолитического метаболизма регулируют активацию дендритных клеток. Кровь (2010) 115:4742–9. doi: 10.1182/blood-2009-10-249540PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
235. Донг Х., Буллок Т.Н. Метаболические влияния, которые регулируют функцию дендритных клеток в опухолях. Фронт Иммунол. (2014) 5:24. doi: 10.3389/fimmu.2014.00024
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
236. Netea-Maier RT, Smit JWA, Netea MG. Метаболические изменения в опухолевых клетках и опухолеассоциированных макрофагах: взаимосвязь. Рак Летт. (2018) 413:102–9. doi: 10.1016/j.canlet.2017.10.037
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
237. Geeraerts X, Bolli E, Fendt SM, Van Ginderachter JA. Метаболизм макрофагов как терапевтическая мишень при раке, атеросклерозе и ожирении.
Фронт Иммунол. (2017) 8:289. doi: 10.3389/fimmu.2017.00289PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
238. Wenes M, Shang M, Di Matteo M, Goveia J, Martin-Perez R, Serneels J, et al. Метаболизм макрофагов контролирует морфогенез кровеносных сосудов опухоли и метастазирование. Сотовый метаб . (2016) 24:701–15. doi: 10.1016/j.cmet.2016.09.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
239. Миллс Э.Л., О’Нил Л.А. Перепрограммирование митохондриального метаболизма в макрофагах как противовоспалительный сигнал. Евро J Иммунол. (2016) 46:13–21. doi: 10.1002/eji.201445427
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
240. Поллард Дж.В. Образованные опухолью макрофаги способствуют прогрессированию опухоли и метастазированию. Nat Rev Cancer (2004) 4:71–8. doi: 10.1038/nrc1256
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
241.
Rotondo R, Barisione G, Mastracci L, Grossi F, Orengo AM, Costa R, et al. IL-8 индуцирует экзоцитоз аргиназы 1 полиморфноядерными нейтрофилами при немелкоклеточном раке легкого. Int J Cancer (2009) 125:887–93. doi: 10.1002/ijc.24448PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
242. Велья Ф., Габрилович Д.И. Дендритные клетки при раке: новый взгляд на роль. Курр Опин Иммунол. (2017) 45:43–51. doi: 10.1016/j.coi.2017.01.002
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
243. Mbongue JC, Nicholas DA, Torrez TW, Kim NS, Firek AF, Langridge WH. Роль индоламин-2,3-диоксигеназы в иммуносупрессии и аутоиммунитете. Вакцины (2015) 3:703–29. doi: 10.3390/vaccines3030703
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
244. Тимосенко Е., Хаджиниколау А.В., Церундоло В. Модуляция канцер-специфических иммунных реакций ферментами, расщепляющими аминокислоты. Иммунотерапия (2017) 9:83–97.
doi: 10.2217/imt-2016-0118PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
245. Godin-Ethier J, Hanafi LA, Piccirillo CA, Lapointe R. Экспрессия индоламин-2,3-диоксигеназы при раке человека: клинические и иммунологические перспективы. Clin Cancer Res. (2011) 17:6985–91. doi: 10.1158/1078-0432.CCR-11-1331
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
246. Мануэль Э.Р., Diamond DJ. Неизведанная дорога, вымощенная IDO глушением: использование противоопухолевой активности нейтрофилов. Oncoimmunology (2013) 2. doi: 10.4161/onci.23322
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
247. Balachandran VP, Cavnar MJ, Zeng S, Bamboat ZM, Ocuin LM, Obaid H, et al. Иматиниб потенцирует противоопухолевый Т-клеточный ответ при гастроинтестинальной стромальной опухоли посредством ингибирования идо. Nat Med . (2011) 17:1094–100. doi: 10.1038/nm.2438
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
248.
фон Бергвельт-Бейлдон М.С., Попов А., Сарич Т., Хемниц Дж., Классен С., Стоффель М.С. и др. CD25 и индоламин-2,3-диоксигеназа активируются простагландином E2 и экспрессируются ассоциированными с опухолью дендритными клетками in vivo : дополнительные механизмы ингибирования Т-клеток. Кровь (2006) 108:228–37. doi: 10.1182/blood-2005-08-3507CrossRef Полный текст | Google Scholar
249. Prendergast GC, Smith C, Thomas S, Mandik-Nayak L, Laury-Kleintop L, Metz R, et al. Индоламин-2,3-диоксигеназные пути патогенного воспаления и иммунного ускользания при раке. Рак Иммунол Иммунотер. (2014) 63:721–35. doi: 10.1007/s00262-014-1549-4
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
250. Hornyák L, Dobos N, Koncz G, Karanyi Z, Páll D, Szabó Z, et al. Роль индоламин-2,3-диоксигеназы в развитии, диагностике и терапии рака. Фронт Иммунол. (2018) 9:151. doi: 10.3389/fimmu.2018.00151
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
251.
Beatty GL, O’Dwyer PJ, Clark J, Shi JG, Bowman KJ, Scherle PA, et al. Первое исследование фазы I на людях перорального ингибитора индоламин-2,3-диоксигеназы-1 эпакадостата (INCB024360) у пациентов с прогрессирующими солидными злокачественными новообразованиями. Клин Рак Рез . (2017) 23:3269–76. doi: 10.1158/1078-0432.CCR-16-2272PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
252. Gangadhar TC, Hamid O, Smith DC, Bauer TM, Wasser JS, Olszanski AJ, et al. Эпакадостат плюс пембролизумаб у пациентов с прогрессирующей меланомой и некоторыми солидными опухолями: обновленные результаты фазы 1 из ECHO-202/KEYNOTE-037. Энн Онкол . (2016) 27 (Приложение 6): 1110PD. doi: 10.1093/annonc/mdw379.06
CrossRef Full Text | Академия Google
253. Holmgaard RB, Zamarin D, Li Y, Gasmi B, Munn DH, Allison JP, et al. Экспрессируемый опухолью IDO рекрутирует и активирует MDSCs Treg-зависимым образом. Представитель ячейки . (2015) 13:412–24.
doi: 10.1016/j.celrep.2015.08.077PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
254. Metz R, Rust S, Duhadaway JB, Mautino MR, Munn DH, Vahanian NN, et al. IDO ингибирует сигнал достаточности триптофана, который стимулирует mTOR: новый эффекторный путь IDO, нацеленный на D-1-метил-триптофан. Онкоиммунология (2012) 1:1460–8. doi: 10.4161/onci.21716
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
255. Sharma MD, Hou DY, Baban B, Koni PA, He Y, Chandler PR, et al. Перепрограммированные регуляторные Т-клетки Foxp3(+) оказывают существенную помощь в поддержке кросс-презентации и примирования CD8(+) Т-клеток у наивных мышей. Иммунитет (2010) 33:942–54. doi: 10.1016/j.immuni.2010.11.022
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
256. Mautino MR, Kumar S, Zhuang H, Waldo J, Jaipuri F, Potturi H, et al. Abstract 4076: новое пролекарство индоксимода с улучшенными фармакокинетическими свойствами.
Исследование рака. (2017) 77 (13 Дополнение): 4076. doi: 10.1158/1538-7445.AM2017-4076CrossRef Full Text | Google Scholar
257. Vasquez-Dunddel D, Pan F, Zeng Q, Gorbounov M, Albesiano E, Fu J, et al. STAT3 регулирует аргиназу-I в супрессорных клетках миелоидного происхождения у больных раком. Дж Клин Инвест . (2013) 123:1580–9. doi: 10.1172/JCI60083
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
258. Ланг Р., Патель Д., Моррис Дж. Дж., Ручман Р. Л., Мюррей П. Дж. Формирование экспрессии генов в активированных и покоящихся первичных макрофагах с помощью IL-10. Дж Иммунол . (2002) 169:2253–63. doi: 10.4049/jimmunol.169.5.2253
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
259. Boutard V, Havouis R, Fouqueray B, Philippe C, Moulinoux JP, Baud L. Трансформирующий фактор роста-бета стимулирует активность аргиназы в макрофагах. Значение для регуляции цитотоксичности макрофагов.
Дж Иммунол . (1995) 155:2077–84.Реферат PubMed | Google Scholar
260. Jost MM, Ninci E, Meder B, Kempf C, Van Royen N, Hua J, et al. Различные эффекты GM-CSF и TGFbeta1 на активность аргиназы-1 макрофагов костного мозга, экспрессию MCP-1 и матриксную металлопротеиназу-12: потенциальная роль во время артериогенеза. Фасеб Дж . (2003) 17:2281–3. doi: 10.1096/fj.03-0071fje
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
261. Corzo CA, Condamine T, Lu L, Cotter MJ, Youn JI, Cheng P, et al. HIF-1альфа регулирует функцию и дифференцировку супрессорных клеток миелоидного происхождения в микроокружении опухоли. J Exp Med . (2010) 207:2439–53. doi: 10.1084/jem.20100587
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
262. de Boniface J, Mao Y, Schmidt-Mende J, Kiessling R, Poschke I. Характер экспрессии иммуномодулирующего фермента аргиназы 1 в крови, лимфатических узлах и опухолевой ткани у пациентов с раком молочной железы на ранней стадии.
Онкоиммунология (2012) 1:1305–12. doi: 10.4161/onci.21678PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
263. Ян Z, Ming XF. Функции изоформ аргиназы в воспалительных реакциях макрофагов: влияние на сердечно-сосудистые заболевания и метаболические нарушения. Фронт Иммунол . (2014) 5:533. doi: 10.3389/fimmu.2014.00533
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
264. Тур С.М., Сайед Хаджа А.С., Эль Салхат Х., Бекдаче О., Канбар Дж., Джалуди М. и др. Повышенный уровень циркулирующих и инфильтрирующих опухоль гранулоцитарных миелоидных клеток у больных колоректальным раком. Фронт Иммунол . (2016) 7:560. doi: 10.3389/fimmu.2016.00560
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
265. Vannini F, Kashfi K, Nath N. Двойная роль iNOS при раке. Редокс Биол . (2015) 6: 334–43. doi: 10.1016/j.redox.2015.08.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
266.
Rodriguez PC, Ochoa AC, Al-Khami AA. Метаболизм аргинина в миелоидных клетках формирует врожденный и адаптивный иммунитет. Фронт Иммунол. (2017) 8:93. doi: 10.3389/fimmu.2017.00093PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
267. Очоа А.С., Зеа А.Х., Эрнандес С., Родригес П.С. Аргиназа, простагландины и супрессорные клетки миелоидного происхождения при почечно-клеточном раке. Clin Cancer Res. (2007) 13 (2 часть 2): 721с-6с. doi: 10.1158/1078-0432.CCR-06-2197
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
268. Rodriguez PC, Quiceno DG, Zabaleta J, Ortiz B, Zea AH, Piazuelo MB, et al. Продукция аргиназы I в микроокружении опухоли зрелыми миелоидными клетками ингибирует экспрессию Т-клеточного рецептора и антиген-специфические Т-клеточные ответы. Рак Res . (2004) 64:5839–49. doi: 10.1158/0008-5472.CAN-04-0465
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
269.
Crittenden MR, Savage T, Cottam B, Baird J, Rodriguez PC, Newell P, et al. Экспрессия аргиназы I в миелоидных клетках ограничивает контроль остаточного заболевания после лучевой терапии опухолей у мышей. Рез. (2014) 182:182–90. doi: 10.1667/RR13493.1PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
270. Бронте В., Касич Т., Гри Г., Галлана К., Борселлино Г., Мариго И. и др. Усиление противоопухолевого ответа Т-лимфоцитов, инфильтрирующих рак предстательной железы человека. J Exp Med. (2005) 201:1257–68. doi: 10.1084/jem.20042028
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
271. Serafini P, Meckel K, Kelso M, Noonan K, Califano J, Koch W, et al. Ингибирование фосфодиэстеразы-5 усиливает эндогенный противоопухолевый иммунитет за счет снижения функции супрессорных клеток миелоидного происхождения. J Exp Med. (2006) 203:2691–702. doi: 10.1084/jem.20061104
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
272.
Meyer C, Sevko A, Ramacher M, Bazhin AV, Falk CS, Osen W, et al. Хроническое воспаление способствует активации клеток-супрессоров миелоидного происхождения, блокирующих противоопухолевый иммунитет в модели меланомы трансгенных мышей. Proc Natl Acad Sci USA. (2011) 108:17111–6. doi: 10.1073/pnas.1108121108PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
273. Lin S, Wang J, Wang L, Wen J, Guo Y, Qiao W, et al. Ингибирование фосфодиэстеразы-5 подавляет онкогенез, вызванный воспалением толстой кишки, путем блокирования рекрутирования MDSC. Am J Cancer Res. (2017) 7:41–52.
Реферат PubMed | Google Scholar
274. De Santo C, Serafini P, Marigo I, Dolcetti L, Bolla M, Del Soldato P, et al. Нитроаспирин корректирует иммунную дисфункцию у носителей опухоли и способствует эрадикации опухоли с помощью противораковой вакцинации. Proc Natl Acad Sci USA . (2005) 102:4185–90. doi: 10.1073/pnas.0409783102
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
275.
Весоловский Р., Марковиц Дж., Карсон В.Е. III. Клетки-супрессоры миелоидного происхождения — новая терапевтическая мишень в лечении рака. J Иммунный рак . (2013) 1:10. doi: 10.1186/2051-1426-1-10PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
276. Califano JA, Khan Z, Noonan KA, Rudraraju L, Zhang Z, Wang H, et al. Тадалафил повышает опухолеспецифический иммунитет у пациентов с плоскоклеточным раком головы и шеи. Клин Рак Рез . (2015) 21:30–8. doi: 10.1158/1078-0432.CCR-14-1716
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
277. Weed DT, Vella JL, Reis IM, De la fuente AC, Gomez C, Sargi Z, et al. Тадалафил снижает количество супрессорных клеток миелоидного происхождения и регуляторных Т-клеток и способствует иммунитету к опухолям у пациентов с плоскоклеточным раком головы и шеи. Clin Cancer Res. (2015) 21:39–48. doi: 10.1158/1078-0432.CCR-14-1711
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
278.
Hassel JC, Jiang H, Bender C, Winkler J, Sevko A, Shevchenko I, et al. Тадалафил обладает биологической активностью в отношении меланомы человека. Результаты пилотного исследования тадалафила у пациентов с метастатической меланомой (TaMe). Онкоиммунология (2017) 6:e1326440. doi: 10.1080/2162402X.2017.1326440PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
279. Kim SH, Roszik J, Grimm EA, Ekmekcioglu S. Влияние метаболизма L-аргинина на иммунный ответ и противораковую иммунотерапию. Онкол передний . (2018) 8:67. doi: 10.3389/fonc.2018.00067
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
280. Talmadge JE, Hood KC, Zobel LC, Shafer LR, Coles M, Toth B. Химиопрофилактика путем ингибирования циклооксигеназы-2 снижает размножение незрелых миелоидных супрессорных клеток. Int Immunopharmacol. (2007) 7:140–51. doi: 10.1016/j.intimp.2006.09.021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
281.
Ерусланов Э., Дауркин И., Ортиз Дж., Виег Дж., Кусмарцев С. Pivotal Advance: опосредованная опухолью индукция миелоидных супрессорных клеток и М2-поляризованных макрофагов путем изменения внутриклеточного катаболизма PGE(2) в миелоидных клетках. . J Лейкок Биол . (2010) 88:839–48. doi: 10.1189/jlb.1209821Полный текст CrossRef | Google Scholar
282. Obermajer N, Muthuswamy R, Lesnock J, Edwards RP, Kalinski P. Положительная обратная связь между PGE2 и COX2 перенаправляет дифференцировку дендритных клеток человека в сторону стабильных супрессорных клеток миелоидного происхождения. Кровь (2011) 118:5498–505. doi: 10.1182/blood-2011-07-365825
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
283. Na YR, Yoon YN, Son DI, Seok SH. Ингибирование циклооксигеназы-2 блокирует дифференцировку макрофагов М2 и подавляет метастазирование в мышиной модели рака молочной железы. ПЛОС ОДИН . (2013) 8:e63451. doi: 10.1371/journal.
pone.0063451PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
284. Chen EP, Markosyan N, Connolly E, Lawson JA, Li X, Grant GR, et al. Делеция миелоидных клеток COX-2 снижает рост опухоли молочной железы за счет усиления функции цитотоксических Т-лимфоцитов. Канцерогенез (2014) 35:1788–97. doi: 10.1093/carcin/bgu053
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
285. Zea AH, Rodriguez PC, Atkins MB, Hernandez C, Signoretti S, Zabaleta J, et al. Миелоидные супрессорные клетки, продуцирующие аргиназу, у больных почечно-клеточным раком: механизм уклонения от опухоли. Рак Res . (2005) 65:3044–8. doi: 10.1158/0008-5472.CAN-04-4505
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
286. Sinha P, Clements VK, Fulton AM, Ostrand-Rosenberg S. Простагландин E2 способствует прогрессированию опухоли, индуцируя супрессорные клетки миелоидного происхождения. Рак Res. (2007) 67:4507–13.
doi: 10.1158/0008-5472.CAN-06-4174PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
287. Veltman JD, Lambers ME, van Nimwegen M, Hendriks RW, Hoogsteden HC, Aerts JG, et al. Ингибирование ЦОГ-2 улучшает иммунотерапию и связано с уменьшением количества супрессорных клеток миелоидного происхождения при мезотелиоме. Целекоксиб влияет на функцию MDSC. BMC Рак (2010) 10:464. doi: 10.1186/1471-2407-10-464
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
288. Iachininoto MG, Nuzzolo ER, Bonanno G, Mariotti A, Procoli A, Locatelli F, et al. Ингибирование циклооксигеназы-2 (ЦОГ-2) ограничивает активность индоламин-2,3-диоксигеназы 1 (IDO1) в клетках острого миелоидного лейкоза. Молекулы (2013) 18:10132–45. doi: 10.3390/molecules180
2PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Вмешательство Screen-Time по снижению веса, ориентированное на детей дома (SWITCH): рандомизированное контролируемое исследование | Международный журнал поведенческого питания и физической активности
- Исследования
- Открытый доступ
- Опубликовано:
- Ральф Мэддисон 1 ,
- Саманта Марш 1 ,
- Louise Foley 2 ,
- Leonard H Epstein 3 ,
- Timothy Olds 4 ,
- Ofa Dewes 5 ,
- Ihirangi Heke 6 ,
- Karen Carter 1 ,
- Яннан Цзян 1 и
- …
- Клиона Ни Мхурчу 1
Международный журнал поведенческого питания и физической активности том 11 , номер статьи: 111 (2014) Процитировать эту статью
8163 Доступ
35 цитирований
6 Альтметрический
Сведения о показателях
Abstract
Исходная информация
Действия, связанные с экраном, такие как просмотр телевизора (ТВ), видеоигры и использование компьютеров, являются обычным малоподвижным поведением среди молодых людей и связаны с повышенным потреблением энергии и избыточным весом.
Предыдущие вмешательства в отношении малоподвижного поведения на дому были ограничены тем, что основное внимание уделялось ребенку, малым размерам выборки и коротким периодам наблюдения. Исследование SWITCH (интервенционное вмешательство для снижения веса с помощью экрана, ориентированное на детей дома) было направлено на определение влияния семейного вмешательства на дому для снижения малоподвижного поведения на основе экрана на состав тела, малоподвижное поведение, физическую активность и диету. более 24 недель у детей с избыточной массой тела и ожирением.Методы
Было проведено параллельное рандомизированное контролируемое исследование с двумя группами. Детей и их основных опекунов, проживающих в Окленде, Новая Зеландия, набирали через школы, общественные центры и из уст в уста. Вмешательство, проводившееся в течение 20 недель, состояло из личной встречи с родителем/опекуном и ребенком для предоставления содержания вмешательства, которое было сосредоточено на обучении и обучении их использованию широкого спектра стратегий, предназначенных для уменьшения экрана их ребенка.
время. Семьям были предоставлены устройства для мониторинга Time Machine TV, которые помогают распределять экранное время, пакеты мероприятий для продвижения альтернативных занятий, онлайн-поддержка через веб-сайт и ежемесячные информационные бюллетени. Участникам контрольной группы был предоставлен интервенционный материал по завершении последующего наблюдения. Первичным результатом было изменение z-показателя ИМТ у детей по сравнению с исходным уровнем до 24 недель.Результаты
Дети (n = 251) в возрасте 9-12 лет и их основной опекун были рандомизированы для получения вмешательства SWITCH (n = 127) или без вмешательства (контрольная группа; n = 124). Не было существенной разницы в изменении zBMI между экспериментальной и контрольной группами, хотя наблюдалась благоприятная тенденция (-0,016; 95% ДИ: -0,084, 0,051; p = 0,64). Значимых различий по вторичным исходам также не было, за исключением тенденции к увеличению физической активности средней интенсивности у детей в группе вмешательства (24,3 мин/д; 9).
5% ДИ: -0,94, 49,51; р = 0,06).Выводы
Семейное вмешательство на дому, направленное на сокращение использования экрана в свободное время, не оказало существенного влияния на время, проводимое перед экраном, или на ИМТ через 24 недели у детей с избыточной массой тела и ожирением в возрасте 9–12 лет.
Регистрация клинических испытаний
Реестр клинических испытаний Австралии и Новой Зеландии
Веб-сайт: http://www.anzctr.org.au
Регистрационный номер исследования: ACTRN12611000164998
Исходная информация
Потребление медиаресурсов детьми и родителями помочь детям удовлетворить образовательные, социальные и развлекательные потребности [1]-[3]. Таким образом, сидячий образ жизни, связанный с экраном (например, просмотр телевизора [ТВ], игра в видеоигры и использование компьютеров), распространен среди молодых людей, при этом 53% новозеландских детей не соблюдают рекомендации по просмотру телевизора менее 2 часов в день.
[4]. Это вызывает беспокойство, учитывая положительную связь между увеличением времени, проведенного за экраном, малоподвижным поведением и неблагоприятными последствиями для здоровья [5].Малоподвижное поведение может быть определено как любое поведение при бодрствовании, характеризующееся низким расходом энергии (т.е. ≤1,5 METS) в сидячем или полулежачем положении [6]. Общее время сидячего образа жизни может включать занятия, связанные с экраном (например, просмотр телевизора или использование компьютера), и занятия, не связанные с экраном (например, чтение, выполнение домашних заданий или использование моторизованного транспорта). Однако с точки зрения последствий для здоровья не все виды малоподвижного образа жизни одинаковы. Например, было показано, что при чтении книги традиционное или нецелевое малоподвижное поведение снижает артериальное давление [7], а малоподвижное поведение на основе экрана оказывает неблагоприятное воздействие на избыточный вес и ожирение [8]–[13].
], метаболический риск [14], внимание [15], просоциальное поведение, самооценка и успеваемость [16].Было проведено большое количество вмешательств, направленных на борьбу с избыточным весом у детей за счет сокращения времени сидячего образа жизни. В то время как систематические обзоры [17]-[24] таких вмешательств, как правило, сообщают о положительных, хотя и небольших размерах эффекта, в ряде этих обзоров участие родителей и ориентация на домашнюю среду отмечены как ключевые аспекты, связанные с успехом вмешательства [21]. [23].
Активное участие родителей может быть особенно важным для обеспечения эффективности экранных вмешательств. Занятия на экране очень приятны и обладают исключительной способностью привлекать и удерживать внимание детей. Родители сообщают, что пользуются этими уникальными возможностями, используя экраны как в качестве электронной няни, так и в качестве образовательного инструмента [2], [25]. Кроме того, физические и межличностные факторы в домашней обстановке связаны с увеличением времени, проводимого детьми перед экраном [26], [27].
Вместе эти факторы представляют собой уникальные барьеры, но также и уникальные возможности для реализации эффективных и устойчивых вмешательств, связанных с малоподвижным образом жизни перед экраном.На сегодняшний день ряд вмешательств в отношении малоподвижного поведения на дому с участием родителей был проведен среди детей и подростков [28]–[35]. Большинство этих вмешательств были сосредоточены на ребенке, а родители в первую очередь были заинтересованы в контроле за использованием различных устройств, таких как телевизионные электронные мониторы времени [28] и активные видеоигры [32], [33]. Кроме того, размеры выборки исследования, как правило, были небольшими, а период наблюдения слишком коротким, чтобы оценить долгосрочный успех таких вмешательств, а выборки были относительно однородными с точки зрения этнической принадлежности [28]. В попытке устранить эти недостатки мы провели большое 6-месячное рандомизированное контролируемое исследование, чтобы изучить эффективность домашнего семейного вмешательства, направленного на снижение малоподвижного поведения на основе экрана и улучшение состава тела у детей с избыточным весом и ожирением.
. Вмешательство было сосредоточено на обучении и обучении основных опекунов использованию широкого спектра стратегий, предназначенных для сокращения времени, проводимого их ребенком перед экраном.Methods
Исследование Screen-Time по снижению веса, ориентированное на детей дома (SWITCH), было параллельным рандомизированным контролируемым исследованием с двумя группами, направленным на предотвращение чрезмерного увеличения веса за счет снижения малоподвижной активности на основе экрана. Испытание проводилось в Новой Зеландии в период с 2010 по 2012 год [36]. Участники имели право на участие, если они были детьми в возрасте 9-12 лет, жили в столичном районе Большого Окленда, использовали электронные средства массовой информации (телевидение, видеоигры, компьютер) не менее 15 часов в неделю, имели избыточный вес или ожирение (согласно Cole International Cut). -баллов) [37], мог говорить и понимать по-английски. Учитывая высокую распространенность ожирения среди маори (коренное население [19%]) и детей тихоокеанских островов (26%) [38], мы стремились набрать равное количество детей маори, тихоокеанских и немаори/нетихоокеанских детей.
Участников попросили самостоятельно выбрать одну этническую группу, с которой они больше всего себя идентифицируют. В Новой Зеландии большинство маори и жителей тихоокеанских островов проживают в районе Большого Окленда, интегрированные в существующие сообщества. Несмотря на то, что между маори и жителями тихоокеанских островов существуют большие различия в состоянии здоровья по сравнению с немаори/нетихоокеанцами, уровни физической активности и занятия в свободное время для этих групп населения одинаковы. В исследовании мог участвовать один ребенок на домохозяйство. Дети были исключены, если у них было какое-либо заболевание, препятствующее регулярной физической активности, или если они жили более чем в одном домохозяйстве и проводили одинаковое время в обоих домохозяйствах. От каждого участвовавшего ребенка также требовалось, чтобы в исследовании принимал участие один основной опекун в возрасте 18 лет и старше, который мог говорить и понимать по-английски. Участники были набраны через школы, общественные центры, церкви, организации первичной медико-санитарной помощи и из уст в уста. Этическое одобрение исследования было получено от Нижнего Южного регионального комитета (LRS/10/09)./039). Письменное информированное согласие было предоставлено опекунами, а письменное согласие было предоставлено детьми.Рандомизация и ослепление
Подходящие участники были рандомизированы в соотношении 1:1 в группу вмешательства или контрольную группу с помощью централизованной компьютерной рандомизации с использованием стратифицированной блокированной рандомизации (с переменными размерами блоков) для поддержания баланса между важными прогностическими факторами. Были рассмотрены два фактора стратификации: пол (мужской и женский) и этническая принадлежность (маори, тихоокеанцы и немаори/не тихоокеанцы). Сокрытие распределения сохранялось до момента рандомизации. Ослепление участников и научных сотрудников было невозможно из-за характера вмешательства.
Процедура
Оценки проводились дома у участников в начале исследования и через 24 недели после рандомизации.
При исходном посещении было получено письменное информированное согласие, а затем были зарегистрированы физические измерения роста, веса, окружности талии и состава тела ребенка. Статус созревания не оценивался в настоящем испытании по причинам логистики и проблем культурной чувствительности. Также оценивались самооценки малоподвижного поведения, физической активности, диеты и удовольствия от малоподвижного поведения и физической активности. Были измерены рост и вес лица, оказывающего первичную медицинскую помощь, а затем был проведен 7-дневный опросник по физической активности. Все измерения были повторены при 24-недельном посещении. Участники обеих групп также получили телефонный звонок через 12 недель, чтобы отслеживать любые нежелательные явления.Вмешательство
Вмешательство было направлено на сокращение малоподвижного поведения детей в свободное время, связанное с просмотром экрана, и проводилось в течение 20 недель. Он был основан на предыдущем испытании, проведенном в Соединенных Штатах (США) Epstein et al.
[28], которые использовали мониторы с ограничением просмотра телевизора, обучение, многократные визиты на дом и стимулы для сокращения просмотра телевизора маленькими детьми (в возрасте 7-9 лет) и использования компьютера. Это исследование было связано с устойчивыми изменениями в течение двух лет в малоподвижном поведении на основе экрана. Содержание интервенции в США было адаптировано для использования в Новой Зеландии и включало более широкий акцент на все действия, связанные с экраном в доме (телевидение, видео и запуск после использования компьютера). Адаптация также включала устранение финансовых стимулов из вмешательства, поскольку это не считалось масштабируемым подходом в Новой Зеландии, изменение содержания, чтобы обеспечить его приемлемость для семей маори (коренных) и тихоокеанских семей, а также включение веб-сайта участников для облегчения доступа к информации. обмен.Содержание вмешательства было основано на социальной когнитивной теории (SCT) [39] и теории поведенческой экономики (BET) [40].
Подробная информация представлена ниже. SCT относится к триадным, взаимным отношениям между личностными, экологическими, поведенческими факторами и окружающей средой [39]. SCT объясняет, как люди приобретают и поддерживают определенные модели поведения, а также обеспечивает основу для стратегий вмешательства [39]. В настоящем вмешательстве мы сосредоточились на внесении изменений в домашнюю и семейную среду, а также на предложении родителям внести личные и поведенческие изменения (изложенные ниже). Согласно BET, выбор заниматься физической активностью включает в себя выбор этого вместо конкурирующего малоподвижного поведения [41], например, выбор ходить в школу пешком, а не водить машину с родителем. Этот выбор зависит от двух факторов: относительной легкости доступа к конкурирующим видам деятельности и их подкрепляющей (наслаждающей) ценности. Ключевой принцип BET заключается в том, что изменение одного поведения приведет к последующим изменениям другого связанного поведения [41]. BET утверждает, что физическая активность и малоподвижный образ жизни связаны как заменители поведения. Следовательно, сокращение времени, проведенного в сидячем положении, может привести к замене физической активностью, поскольку это время перераспределяется [41]. Однако сила обратной зависимости между физической активностью и малоподвижным поведением («перекрестная эластичность использования времени») зависит от конкретных видов деятельности и рассматриваемых временных блоков. Более того, полной замены малоподвижного образа жизни физической активностью от умеренной до высокой, скорее всего, не произойдет; одно малоподвижное поведение (например, просмотр телевизора) может быть заменено другим (например, чтением) или физической активностью легкой интенсивности. Интервенционные исследования, проведенные у детей, показывают, что снижение целевого малоподвижного поведения (время перед экраном) было связано со значительным увеличением физической активности, хотя нецелевое малоподвижное поведение также увеличилось [42]-[45]. Основное внимание уделялось обучению основных опекунов тому, как инициировать изменения в домашней обстановке, чтобы облегчить изменение поведения ребенка и реализовать стратегии изменения поведения. Семьям были предложены три элемента: (1) предоставление стратегий изменения поведения, (2) помощь в бюджетировании времени в СМИ и (3) пакет мероприятий для детей.Предоставление стратегий изменения поведения
Воспитателям первичного звена было предложено обучение и поддержка для реализации стратегий в домашних условиях по сокращению использования средств массовой информации. Хотя в рамках исследования SWITCH оценивался только один ребенок на домохозяйство, вмешательство было сосредоточено на всей домашней обстановке (в соответствии с SCT). Во время первой личной встречи обученные ассистенты-исследователи предоставили культурно значимое образование основному опекуну. Ролевое моделирование и обучение через наблюдение являются ключевыми принципами SCT, а подкрепляющая ценность является ключевым фактором, определяющим выбор поведения согласно BET. Таким образом, вмешательство было направлено на изменение обоих этих детерминантов, чтобы сократить время, проводимое за экраном.
Во время личной встречи лицам, осуществляющим уход, был предоставлен обзор вмешательства, и им было предложено включить похвалу, положительное подкрепление, составление бюджета контроля окружающей среды и самоконтроль, позитивное ролевое моделирование и предложить альтернативные виды деятельности на основе экранных средств массовой информации. Семьи также получали краткие ежемесячные информационные бюллетени с изложением дополнительных стратегий по сокращению малоподвижной активности с использованием экрана, которые включали описание и обоснование использования соответствующих стратегий, а также практические советы по их реализации. Различные версии каждого информационного бюллетеня были доступны для семей маори, тихоокеанских и немаори/нетихоокеанских семей. Хотя содержание информационных бюллетеней было одинаковым для каждой группы, визуальное представление и язык различались. Защищенный веб-сайт был также предоставлен для поддержки содержания вмешательства и включал ежемесячные информационные бюллетени в электронном формате, дополнительные советы и информацию о том, как уменьшить активность, связанную с экраном, и альтернативные варианты (такие как настольные игры, выполнение домашних заданий, искусство и ремесла, использование одного предметов из пакета действий SWITCH [см. ниже], игры на свежем воздухе и физическая активность) и ссылки на программы деятельности сообщества [34].Бюджетирование использования СМИ
На основании предыдущей экспериментальной работы [34], семьи в группе вмешательства получили по два устройства ТВ-мониторинга Time Machine (Family Safe Media, Park City, США) для помощи в составлении бюджета на использование средств массовой информации. Time Machine была подключена к телевизору или другому мультимедийному устройству (например, DVD-плееру, игровой приставке), но подключить устройство к компьютеру не удалось. Кабели Time Machine были защищены и заблокированы, чтобы предотвратить несанкционированный доступ, а родители получили ключи для доступа к кабелям в случае необходимости. Каждая машина времени поставлялась с 30 токенами, каждый из которых позволял просматривать 30 минут; однако опекуны могли распределять их по своему усмотрению.
Набор для занятий
Детям в группе вмешательства был выдан набор для занятий, содержащий предложения и варианты занятий без использования экрана, включая цветные карандаши, отрезок веревки, игральные карты, теннисный мяч для игры в гандбол или подобные игры, и карточки действий, которые описывали простые игры или занятия, в которые можно играть.
Дальнейшая адаптация вмешательства также включала предложение детям настольных игр, характерных для маори, и инструкций по традиционным играм маори.Точность проведения вмешательства
Проведен мониторинг проведения вмешательства общественными работниками. Член исследовательской группы посетил личное посещение и наблюдал за каждым из социальных работников, предоставляющих содержание вмешательства, и зафиксировал отсутствие или наличие обсуждаемых методов, используя стандартный формат. В конце оценки исследователь предоставил обратную связь социальному работнику, чтобы убедиться, что все компоненты вмешательства были реализованы.
Контрольная группа
Контрольной группе было предложено вести себя как обычно, и она получила доступ к общему общедоступному веб-сайту SWITCH. Через 12 недель с основным опекуном в обеих группах связались для подтверждения контактной информации и регистрации любых серьезных нежелательных явлений. По окончании наблюдения контрольной группе были предложены компоненты вмешательства.
Исходы
Все исходы оценивались на исходном уровне и через 24 недели.
Основной результат
Первичным результатом было изменение z-показателя ИМТ детей (стандартизированного по возрасту и полу с использованием стандарта роста ВОЗ 2007 г.) от исходного уровня до 24 недель.
Вторичные исходы
Ребенок: ИМТ (кг/м 2 ), масса тела (кг), окружность талии (см), процентное содержание телесного жира (%), самооценка показателей ежедневной физической активности (PA; включая минуты) общей ФА, ФА легкой интенсивности [LPA] и ФА от умеренной до энергичной [MVPA]), общее время сидячего образа жизни (минуты), сон, потребление пищи (ежедневное потребление энергии [кДж], энергия, потребляемая с закусками, и частота легких употребление алкоголя) и воспринимаемое удовольствие от физической активности и малоподвижного образа жизни.
Основной опекун: ИМТ и самооценка физической активности (ежедневные минуты общего PA, LPA и MVPA).
Измерения
Антропометрия
Антропометрические измерения проводились согласно стандартным методикам [46].
ИМТ рассчитывали на основе данных о росте и весе (кг/м 2 ) и преобразовывали в стандартизированный z-показатель с использованием возрастных и половых эталонов роста ВОЗ 2007 г. для 5–19 лет (www.who.int/growthref). Состав тела оценивали по биоимпедансу на приборе ImpediMed DF50 Bioimpedence Monitor (Квинсленд, Австралия). Свободная от жира масса (FFM), жировая масса (FM) и процент жира в организме были рассчитаны для всех участников с использованием специфических для Новой Зеландии уравнений [47].Физическая активность и малоподвижный образ жизни
Физическая активность детей и малоподвижный образ жизни измерялись с помощью мультимедийной системы воспоминаний об активности для детей и подростков (MARCA) [48], компьютеризированного самоотчета, 24-часового напоминания об использовании времени, которое использует формат сегментированного дня с самоопределяемыми опорными точками. MARCA имеет надежность повторного тестирования в тот же день r = 0,84–0,92 для основных переменных исхода и достоверность относительно акселерометрии r = 0,45 для уровня физической активности (PAL) [48].
Детей просили вспомнить, чем они занимались в течение двух предыдущих дней (48 часов). Когда ребенок сообщал об участии в двух видах деятельности одновременно, для целей анализа использовалась следующая иерархия: физическая активность, экранное время, активный транспорт, пассивный транспорт и «другие» виды деятельности. Например, если ребенок сообщил, что смотрит DVD одновременно с тем, что едет куда-то, это действие будет засчитываться как экранное время. Время, затраченное на каждое действие, суммировалось, чтобы определить, сколько времени каждый участник провел в общей PA, LPA (≥ 1,5 MET и <3 MET), MPA (≥ 3 MET и <6 MET), активной физической активности (VPA; ≥ 6 MET). ), передвижение, общее время сидячего образа жизни (активность вне сна <3 MET), время сидячего образа жизни на основе экрана, время сидячего образа жизни без экрана и сон.Физическая активность основного лица, осуществляющего уход, измерялась с использованием полной формы Международного вопросника физической активности (IPAQ-LF) [49], который оценивает физическую активность в конкретных областях, ходьбу и сидение.
MET-минуты в неделю рассчитывались как продолжительность × частота в неделю × значение MET, которое суммировалось по областям активности для получения оценки общей PA по всем сообщаемым видам деятельности.Потребление пищи
Полуколичественный вопросник частоты приема пищи (FFQ) использовался для регистрации информации о потреблении пищи. FFQ был разработан для исследования детского питания в Новой Зеландии [50] и имеет такую же воспроизводимость, что и FFQ для детей и подростков, используемые в других странах [51]. В него вошли 104 наиболее часто употребляемых продукта питания. Детей спрашивали, как часто и сколько они ели каждого из продуктов за предыдущие четыре недели. Путем использования стандартизированных размеров порций было оценено общее ежедневное потребление энергии, а также потребление энергии из закусок. Также регистрировалось потребление определенных продуктов питания (например, закусок и подслащенных сахаром напитков).
Удовольствие от физической активности и малоподвижного образа жизни
Были измерены психологические переменные для определения их потенциального опосредующего эффекта.
Воспринимаемое удовольствие от физической активности оценивали с помощью Шкалы удовольствия от физической активности из 14 пунктов [52], адаптированной для использования подростками в 2001 году [53]. Воспринимаемое удовольствие от малоподвижного образа жизни оценивали с использованием шкалы, адаптированной из Salmon et al. [54].Оценка процесса
Основные опекуны, участвующие в вмешательстве, заполнили анкету на выходе, чтобы определить их восприятие вмешательства и использование ими компонентов вмешательства.
Размер выборки
Априорная оценка размера выборки из показала, что 270 детей (135 в каждой группе) обеспечат мощность не менее 90% при уровне значимости 5% (двусторонняя) для обнаружения разницы в 0,2 единицы в изменении zBMI от исходного уровня до 24 недель между экспериментальной и контрольной группами при стандартном отклонении 0,5. Выборка также обеспечит 80% мощности для обнаружения разницы в 0,3 единицы в изменении zBMI между этническими группами (маори, тихоокеанцы и немаори/не тихоокеанцы).
Статистический анализ
Формальный план статистического анализа (SAP) был одобрен Руководящим комитетом исследования до блокировки данных. Статистический анализ был выполнен с использованием SAS версии 9.3 (SAS Institute Inc. Cary NC) и R версии 2.15 (R Foundations for Statistical Computing). Все статистические тесты были двусторонними, и на протяжении всего анализа сохранялся уровень значимости 5%. Оценка лечения проводилась по принципу намерения лечить (ITT) с использованием данных, собранных у всех рандомизированных участников. К отсутствующим данным (если таковые имеются) для основного результата был применен метод множественного вменения. Вменение вторичных исходов не проводилось. Модели регрессии анализа ковариации (ANCOVA) использовались для оценки основных эффектов лечения на первичные и вторичные исходы через 24 недели с поправкой на исходное значение исхода, возраст, пол и этническую принадлежность.
Результаты
Четыреста четыре ребенка зарегистрировались для участия в исследовании SWITCH, из них 251 (62%) имели право участвовать.
Таблица 1 участников исследования Дети были случайным образом распределены в экспериментальную (n = 127) и контрольную (n = 124) группы, при этом 121 (95%) и 117 (94%) завершили 24-недельное наблюдение. Большинство детей были мужского пола (57%) тихоокеанского происхождения (n = 133, 53%), средний возраст 11 лет. Группы лечения были хорошо сбалансированы по исходным характеристикам (таблица 1).Полноразмерная таблица
Первичный результат
Через 24 недели средние изменения ИМТ по сравнению с исходным уровнем составили 0,03 и 0,05 в экспериментальной и контрольной группах, соответственно, с разницей между группами -0,01 (95% ДИ -0,08). до 0,05), что не было статистически значимым (р = 0,64). Результаты были схожими с использованием множественных импутаций для небольшой доли отсутствующих данных с групповой разницей -0,01 (95% ДИ от -0,08 до 0,05; p = 0,67).
Вторичные исходы
Антропометрия
Не было никаких существенных изменений в ИМТ, окружности талии, безжировой массе, жировой массе, процентном соотношении телесного жира или ИМТ родителей/опекунов между экспериментальной и контрольной группами через 24 недели (таблица 2).
Таблица 2 Различия в лечении лиц, осуществляющих основной уход, через 24 недели (вмешательство – контроль)Полноразмерный стол
Физическая активность, малоподвижный образ жизни и сон
В конце 24-недельного вмешательства не было выявлено существенных различий в показателях физической активности, малоподвижного поведения или сна между экспериментальной и контрольной группами. Однако через 24 недели в группе вмешательства наблюдалась тенденция к увеличению МПА по сравнению с исходным уровнем (+24,3; 95% ДИ: -0,94, 49,51; p = 0,06). В 24 недели дети как в экспериментальной, так и в контрольной группах проводили большую часть своего времени в малоподвижном образе жизни (примерно 8,9 часов в день). Хотя это и не является статистически значимым, обе группы сообщили об уменьшении продолжительности малоподвижного образа жизни через 24 недели (таблица 1).
Учитывая, что вмешательство было направлено на модификацию экранных занятий в свободное время, был проведен анализ чувствительности для определения влияния вмешательства на сидячий образ жизни (экранный и неэкранный) после школы и уровни физической активности.
Тот же регрессионный анализ был проведен для изучения данных об использовании времени в будние дни с 15:00 до 18:00, а также в выходные дни; однако существенных различий не наблюдалось.Удовольствие детей от физической активности и сидячий образ жизни существенно не отличались после вмешательства.
Потребление пищи
После 24-й недели не было различий в потреблении пищи, о котором сообщали сами пациенты, между экспериментальной и контрольной группами.
Основной опекун
После 24-недельного вмешательства не было выявлено существенных различий между экспериментальной и контрольной группами по ИМТ, физической активности или малоподвижному поведению лиц, осуществляющих уход (таблица 2).
Оценка процесса
Почти половина (46%) участников сообщили, что никогда не использовали Time Machine для составления бюджета на использование телевизора или компьютера их ребенком; однако 57% сообщили об использовании любой из стратегий, обсуждаемых в ежемесячных информационных бюллетенях.
На прошлой неделе 43% лиц, осуществляющих уход, сообщили об использовании какой-либо из стратегий, позволяющих изменить время от времени до частого использования экрана.Обсуждение
24-недельное семейное вмешательство на дому не оказало существенного влияния на ИМТ у детей в возрасте от 9 до 12 лет в Новой Зеландии. Кроме того, вмешательство не было связано со значительными изменениями в физической активности, малоподвижном образе жизни, сне, рационе питания или предпочтениях в отношении физической активности и малоподвижного образа жизни. Вмешательство также не оказало никакого влияния на ИМТ, физическую активность, малоподвижное поведение и предпочтения лиц, осуществляющих уход. Несмотря на эти нулевые результаты, наблюдались устойчивые эффекты тенденции для большинства представляющих интерес показателей, о которых сообщали сами пациенты, включая снижение общего времени сидячего образа жизни, времени сидячего образа жизни на основе экрана и общего потребления энергии, а также увеличение MVPA.
В совокупности это говорит о том, что вмешательство было недостаточно интенсивным, чтобы манипулировать этим поведением и положительно повлиять на массу тела.Сильные стороны и ограничения
Сильные стороны исследования включали рандомизированный дизайн, сбор данных о физической активности ребенка и его основного опекуна, использование инструментов измерения (MARCA и FFQ), которые ранее были проверены в аналогичных популяциях, адекватный размер выборки для обнаружения изменения ИМТ и относительно длительного наблюдения. Кроме того, наше вмешательство было сосредоточено в первую очередь на сокращении времени сидячего образа жизни, а не на включении других сообщений о здоровье, и использовало устройство блокировки телевизора в дополнение к нескольким стратегиям модификации поведения, две стратегии, которые недавно были определены как важные функции вмешательства, связанные с большим сокращением малоподвижного образа жизни. экранного поведения у детей [22]. Наконец, по сравнению с аналогичными испытаниями [28], наша выборка была более гетерогенной с точки зрения этнической принадлежности и включала значительное число детей тихоокеанских островов и коренных народов маори, которые больше всего страдают от ожирения [4].
Ограничением этого исследования были самостоятельные измерения времени сидячего образа жизни и физической активности. В предыдущих исследованиях использовались более объективные меры, в том числе телевизионные мониторы для захвата экранного времени [28]; однако по логистическим причинам в нашем испытании это было невозможно. В настоящем исследовании, несмотря на то, что мы использовали надежный и действующий инструмент припоминания об использовании времени [48], его чувствительность к оценке изменений менее ясна. Во-вторых, вмешательство не оказало ожидаемого влияния на целевое поведение (сидячий образ жизни и физическая активность), поэтому отсутствие эффекта на массу тела неудивительно. Это отсутствие эффекта может быть объяснено предвзятостью, связанной с показателями самоотчета, недостаточной интенсивностью, связанной с вмешательством, и/или плохим соблюдением. Вмешательство было проведено на разовой встрече с основным опекуном, с которым снова связались только через 12 недель, чтобы подтвердить контактные данные и отследить неблагоприятные события.
Возможно, этого времени было недостаточно, чтобы вызвать изменения в ребенке. Наконец, были также некоторые технические проблемы, связанные с тем, что Time Machine не работала со спутниковым или платным телевидением или с бытовыми компьютерами, что могло ограничивать возможности устройства для помощи в семейном бюджете использования экрана.В контексте предыдущей работы
Хотя настоящее исследование было смоделировано по образцу предыдущей работы Epstein et al. [28] наши результаты значительно различались. Несоответствие в наших выводах можно объяснить следующим. Во-первых, наша старшая выборка детей (9-12 лет) может быть менее податлива родительскому влиянию по сравнению с младшей выборкой (4-7 лет) в исследовании Эпштейна. Согласно недавнему систематическому обзору вмешательств, ориентированных на семью, направленных на снижение малоподвижного образа жизни [55], вмешательства, направленные на детей младшего возраста, могут быть более эффективными для сокращения времени сидячего образа жизни, чем вмешательства, направленные на детей старшего возраста.
Во-вторых, мы использовали другой монитор времени (Time Machine и TV Allowance) для планирования экранного времени из-за логистических проблем, связанных с разницей в напряжении между двумя странами. Хотя в нашем пилотном исследовании предполагалось, что участники будут использовать такую технологию для изменения экранного времени [34], данные выходных интервью показали, что это не так. В-третьих, мы не использовали финансовые стимулы для изменения поведения, поскольку это не считалось масштабируемым вариантом доставки в рамках текущей программы или на уровне населения. Хотя использование стимулов для воздействия на изменение поведения может работать в краткосрочной перспективе, за исключением устойчивых эффектов (2 года), наблюдаемых Эпштейном и соавт. [28] мало доказательств того, что стимулы способствуют долгосрочным изменениям в поведении [56]. С точки зрения поведенческой экономики удаление финансового стимула из вмешательства может быть особенно важным при объяснении различий в результатах между нашим исследованием и исследованием, проведенным Эпштейном и соавт. [28]. Исследования показали, что дети используют средства массовой информации на основе экрана, потому что они высоко ценятся и вознаграждаются [57], поэтому попытки заменить их нефинансовыми вознаграждениями, такими как настольные игры, игры с братьями и сестрами или увеличение физической активности, могут быть менее ценными. и поэтому менее успешны. В целом, несмотря на то, что содержание этого вмешательства было изменено по сравнению с исследованием в США [28], несоблюдение точности первоначальной программы могло объяснить невозможность воспроизвести результаты.В соответствии с Epstein et al. [28] наше вмешательство было разработано для нацеливания на аспекты SCT и BET. Например, вмешательство побуждало родителей/опекунов выступать в качестве ролевых моделей для сокращения времени, проводимого перед экраном. Однако это могло оказаться трудным, если родители также высоко ценили экранное время, такое как просмотр телевизора. Кроме того, во время личной встречи и в информационных бюллетенях лицам, осуществляющим уход, было предложено включить похвалу, положительное подкрепление, составление бюджета контроля окружающей среды и самоконтроль, а также предложить альтернативные виды деятельности на основе экранных средств массовой информации.
Хотя это ключевые конструкции BET, нам неясно, в какой степени родители/опекуны применяли эти методы. Выездные опросы проводились, но более интенсивный мониторинг их проведения не проводился по двум причинам. Во-первых, SWITCH было прагматичным испытанием, в котором исследование было разработано для максимизации экологической достоверности результатов [58], а более интенсивный мониторинг мог быть вмешательством сам по себе. Во-вторых, ресурсы, необходимые для такого интенсивного мониторинга, не входили в рамки этого испытания. Тем не менее, мы предприняли контрольные визиты, чтобы определить достоверность выполнения вмешательства социальными работниками.Хотя было показано, что ряд вмешательств на дому значительно сокращает время, проводимое детьми перед экраном, эти исследования, как правило, проводились на небольших выборках, а долгосрочная эффективность не оценивалась (за исключением Epstein et al. [28]). По сравнению со многими предыдущими исследованиями, несмотря на то, что SWITCH не продемонстрировал значительного влияния на сокращение экранного времени, его последующее наблюдение было более длительным, а размер выборки значительно больше.
На самом деле, подобные нулевые эффекты на время, проводимое перед экраном, также были продемонстрированы в других домашних вмешательствах, в которых использовались большие размеры выборки и более длительные периоды наблюдения [30,32].SWITCH — одно из немногих исследований, посвященных изучению влияния вмешательства на исходы как для ребенка, так и для лиц, осуществляющих уход. Появляется все больше данных, подтверждающих важность привлечения лиц, осуществляющих уход, при проведении мероприятий по борьбе с детским ожирением, поскольку поведение лиц, осуществляющих уход, влияет на поведение ребенка либо напрямую, через совместное поведение (например, совместный просмотр телевизора), либо косвенно, через общую среду [59]. Учитывая важность влияния родителей/опекунов на домашнюю обстановку (например, установление правил/рекомендаций по ограничению времени просмотра домашних экранов, влияние на физическую среду и покупку/наличие домашних продуктов), неудивительно, что поведение детей не изменилось.
когда не наблюдалось изменений в поведении их опекунов или ИМТ.Будущие направления и последствия
Появляется все больше данных, подтверждающих активное воздействие на лиц, осуществляющих уход, при вмешательстве в малоподвижный образ жизни; однако уровень интенсивности ухода, по-видимому, является важным фактором, определяющим успех вмешательства [55]. Таким образом, несмотря на то, что это испытание было отрицательным, SWITCH можно рассматривать как урок по плану вмешательства. В будущих исследованиях, возможно, потребуется увеличить время контакта с опекунами, чтобы помочь им добиться изменений на уровне семьи. Кроме того, точное повторение исследования Epstein et al. [28] протокол может дать положительные результаты.
Хотя родители признают, что дети проводят много времени, взаимодействуя с экранами, они также могут не ограничивать время, проводимое за экраном, опасаясь негативного влияния на развитие своего ребенка. Исследования показывают, что родители часто называют просмотр телевизора и использование компьютера важными средствами обучения [2].
Кроме того, опекуны могут недооценивать серьезность воздействия средств массовой информации на детей, поскольку высокий уровень малоподвижного поведения перед экраном теперь может быть нормализован в обществе; они также могут недооценивать влияние такого поведения на результаты в отношении здоровья. Поэтому в будущих исследованиях следует оценить восприятие лицами, осуществляющими уход, времени, проведенного перед экраном, и его влияние на здоровье, чтобы обосновать будущие вмешательства.Было высказано предположение, что сократить время сидячего образа жизни перед экраном может быть легче, чем увеличить физическую активность [16]; тем не менее, все более очевидной становится ограниченная способность вмешательств эффективно сокращать время, проводимое за компьютером в сидячем положении. Поведение на основе экрана очень полезно. Недавние достижения в области миниатюризации и портативности экранов, доступность контента, предназначенного специально для молодежи, а также повсеместное распространение экранов (iPad, iPod и т.
д.) означают, что возможности экранного воздействия выросли в геометрической прогрессии [60]. Таким образом, ограничение или бюджетирование доступа на уровне домохозяйств становится все труднее, как показано в настоящем исследовании.Выводы
Домашнее семейное вмешательство, направленное на сокращение использования экрана в свободное время, не оказало существенного влияния на время, проводимое перед экраном, или на ИМТ у детей в возрасте 9–12 лет с избыточным весом и ожирением.
Вклад авторов
RM инициировал испытание и обеспечил надзор за исследованием. SM провела исследование и провела сбор данных. KC обеспечивал общую координацию судебного разбирательства. YJ провел статистический анализ. RM, LF, CNM, LHE, TO, OD, IH разработали исследование (концепция проекта и разработка общего плана исследования). RM и SM написали статью. Все авторы прочитали и одобрили окончательный вариант рукописи. RM берет на себя основную ответственность за окончательный контент.
RM имеет полный доступ ко всем данным исследования и берет на себя ответственность за целостность данных и точность анализа данных. Все авторы прочитали и одобрили окончательный вариант рукописи.Сокращения
- ПЕРЕКЛЮЧАТЕЛЬ:
Вмешательство Screen-Time по снижению веса, предназначенное для детей в домашних условиях
- ТВ:
Телевидение
- ИМТ:
Индекс массы тела
- МЕТС:
Метаболические эквиваленты
- IPAQ-LF:
Полная форма международного вопросника физической активности
- FFQ:
Опросник частоты приема пищи
- ЖЖМ:
Обезжиренная масса
- FM:
Жировая масса
- Марка:
Отзыв мультимедийных занятий для детей и подростков
- SCT:
Теория социального познания
- СТАВКА:
Теория поведенческой экономики
- PAL:
Уровень физической активности
- МПУ:
Легкая физическая активность
- МВПА:
Физическая активность от умеренной до высокой интенсивности
- ВПА:
Энергичный физический
Ссылки
«>
Хе М., Ирвин Д.Д., Сангстер Бук Л.М., Такер П., Поллетт Г.Л.: Поведение дошкольников при просмотре экрана: Восприятие родителей. Am J Prev Med. 2005, 29(2): 120-125. 10.1016/j.amepre.2005.04.004.
Артикул Google ученый
Rideout VJ, Hamel E: Медиа-семья: электронные медиа в жизни младенцев, малышей, дошкольников и их родителей. 2006, Семейный фонд Генри Дж. Кайзера, Менло-Парк, Калифорния
Google ученый
Валкенбург П.М., Питер Дж.: Социальные последствия Интернета для подростков: десятилетнее исследование. Curr Dir Psychol Sci. 2009 г., 18 (1): 1-5. 10.1111/j.1467-8721.2009.01595.х.
Артикул Google ученый
Здоровье новозеландских детей: основные результаты исследования здоровья Новой Зеландии 2011/12. 2012, Веллингтон, Министерство здравоохранения
Network SBR: Письмо в редакцию: стандартизированное использование терминов «сидячий образ жизни» и «сидячий образ жизни». Appl Physiol Nutr Metab. 2012, 37 (3): 540-542. 10.1139/х3012-024.
Артикул Google ученый
Гопинат Б., Баур Л., Харди Л., Кифли А., Роуз К., Вонг Т., Митчелл П.: Связь между малоподвижным образом жизни и кровяным давлением в раннем подростковом возрасте. Дж. Гум Гипертенс. 2011, 26 (6): 350-356. 10.1038/jhh.2011.40.
Артикул Google ученый
Dietz WH, Gortmaker SL: Откармливаем ли мы наших детей у телевизора? Ожирение и телесмотрение у детей и подростков.
Педиатрия. 1985, 75 (5): 807-812.Google ученый
Клеланд В.Дж., Шмидт М.Д., Дуайер Т., Венн А.Дж.: Просмотр телевизора и абдоминальное ожирение у молодых людей: связь опосредована потреблением пищи и напитков во время просмотра или снижением физической активности в свободное время?. Am J Clin Nutr. 2008, 87 (5): 1148-1155.
КАС Google ученый
Гортмейкер С.Л., Муст А., Соболь А.М., Петерсон К., Колдитц Г.А., Дитц В.Х. Просмотр телевидения как причина растущего ожирения среди детей в США, 1986-1990 гг. Arch Pediatr Adolesc Med. 1996, 150 (4): 356-362. 10.1001/archpedi.1996.021702
003.
КАС Статья Google ученый
Элгар Ф.Дж., Робертс С., Мур Л., Тюдор-Смит С. Малоподвижный образ жизни, физическая активность и проблемы с весом у подростков в Уэльсе.
Паб Здоровье. 2005, 119(6): 518-524. 10.1016/j.puhe.2004.10.011.КАС Статья Google ученый
Робинсон, штат Теннесси: Сокращение просмотра детьми телевизора для предотвращения ожирения: рандомизированное контролируемое исследование. ДЖАМА. 1999, 282 (16): 1561-1567. 10.1001/jama.282.16.1561.
КАС Статья Google ученый
Рей-Лопес Дж.П., Висенте-Родригес Г., Биоска М., Морено Л.А. Малоподвижное поведение и развитие ожирения у детей и подростков. Nutr Metab Cardiovasc Dis. 2008, 18 (3): 242-251. 10.1016/j.numecd.2007.07.008.
Артикул Google ученый
Ekelund U, Brage S, Froberg K, Harro M, Anderssen SA, Sardinha LB, Riddoch C, Andersen LB: Просмотр телевизора и физическая активность независимо связаны с метаболическим риском у детей: Европейское исследование сердца молодежи.
PLoS Med / Public Library Sci. 2006, 3 (12): e488.Google ученый
Кристакис Д.А., Циммерман Ф.Дж., ДиДжузеппе Д.Л., Маккарти К.А.: Раннее телевизионное воздействие и последующие проблемы с вниманием у детей. Педиатрия. 2004, 113 (4): 708-713. 10.1542/педс.113.4.708.
Артикул Google ученый
Tremblay MS, LeBlanc AG, Kho ME, Saunders TJ, Laouche R, Colley RC, Goldfield G, Gorber SC: Систематический обзор малоподвижного поведения и показателей здоровья у детей школьного возраста и молодежи. Int J Behav Nutr Phys. 2011, 8: 98-10.1186/1479-5868-8-98.
Артикул Google ученый
Biddle SJ, O’Connell S, Braithwaite RE: Вмешательства в малоподвижный образ жизни у молодых людей: метаанализ. Br J Sports Med. 2011, 45 (11): 937-942. 10.1136/bjsports-2011-0.
Артикул Google ученый
DeMattia L, Lemont L, Meurer L: Изменяют ли меры по ограничению малоподвижного образа жизни поведение и снижают лишний вес у детей? Критический обзор литературы. Обес Рев. 2007, 8 (1): 69-81. 10.1111/j.1467-789X.2006.00259.x.
КАС Статья Google ученый
Леунг М.М., Агаронов А., Гриценко К., Йех М.-К.: Меры по снижению малоподвижного образа жизни и детского ожирения среди молодежи школьного возраста: систематический обзор рандомизированных исследований. Дж. Обес. 2012, 2012: 685430-10.1155/2012/685430.
Артикул Google ученый
Maniccia DM, Davison KK, Marshall SJ, Manganello JA, Dennison BA: Метаанализ вмешательств, направленных на сокращение экранного времени детей. Педиатрия. 2011, 128 (1): e193-e210.
10.1542/пед.2010-2353.Артикул Google ученый
Шмидт М.Е. С., Хейнс Дж., О’Брайен А., Макдональд Дж., Прайс С., Шерри Б., Таверас Э.М.: Систематический обзор эффективных стратегий по сокращению времени, проводимого перед экраном у маленьких детей. Ожирение. 2012, 20 (7): 1338-1354. 10.1038/обык.2011.348.
Артикул Google ученый
Стивс Дж.А., Томпсон Д.Л., Бассет Д.Р., Фитцхью Э.К., Рейнор Х.А., Стивс Дж.А., Томпсон Д.Л., Бассетт Д.Р., Фитцхью Э.К., Рейнор Х.А.: Обзор различных стратегий модификации поведения, разработанных для уменьшения малоподвижного поведения перед экраном у детей. Дж. Обес. 2012, 2012: 16-10.1155/2012/379215.
Артикул Google ученый
Van Grieken A, Ezendam NPM, Paulis WD, van der Wouden JC, Raat H: Первичная профилактика избыточного веса у детей и подростков: метаанализ эффективности вмешательств, направленных на снижение малоподвижного образа жизни.
Int J Behav Nutr Phys Act. 2012, 9(1): 61-10.1186/1479-5868-9-61.Артикул Google ученый
Wahi G, Parkin PC, Beyene J, Uleryk EM, Birken CS: Эффективность вмешательств, направленных на сокращение экранного времени у детей: систематический обзор и метаанализ рандомизированных контролируемых исследований. Arch Pediatr Adolesc Med. 2011, 165 (11): 979-986. 10.1001/архопедиатрия.2011.122.
Артикул Google ученый
Коскинен П., Уилсон Р., Йенсема К. Телевидение с субтитрами: новый инструмент для обучения чтению. Читающий мир. 1985, 24 (4): 1-7. 10.1080/19388078509557842.
Артикул Google ученый
Саэленс Б.Е., Саллис Дж.Ф., Надер П.Р., Бройлс С.Л., Берри К.С., Тарас Х.Л.: Домашняя окружающая среда влияет на то, как дети смотрят телевизор с раннего до среднего детства.
J Develop Behav Ped. 2002, 23 (3): 127-132. 10.1097/00004703-200206000-00001.Артикул Google ученый
Салмон Дж., Тимперио А., Телфорд А., Карвер А., Кроуфорд Д.: Ассоциация семейного окружения с просмотром детских телепередач и низким уровнем физической активности. Ожирение. 2005, 13 (11): 1939-1951. 10.1038/обык.2005.239.
Артикул Google ученый
Эпштейн Л.Х., Реммих Дж.Н., Робинсон Дж.Л., Палух Р.А., Виневич Д.Д., Фюрх Дж.Х., Робинсон Т.Н.: Рандомизированное исследование влияния сокращения просмотра телевизора и использования компьютера на индекс массы тела у детей раннего возраста. Arch Pediatr Adolesc Med. 2008, 162 (3): 239-245. 10.1001/архопедиатрия.2007.45.
Артикул Google ученый
Вера М.С., Берман Н., Хео М., Пьетробелли А.
, Галлахер Д., Эпштейн Л.Х., Эйден М.Т., Эллисон Д.Б.: Влияние условного телевидения на физическую активность и просмотр телепередач у детей с ожирением. Педиатрия. 2001, 107 (5): 1043-1048. 10.1542/пед.107.5.1043.КАС Статья Google ученый
French SA, Gerlach AF, Mitchell NR, Hannan PJ, Welsh EM: Профилактика ожирения в домашних условиях: примите меры — групповое рандомизированное исследование. Ожирение. 2011, 19 (10): 2082-2088. 10.1038/обык.2010.328.
Артикул Google ученый
Goldfield GS, Mallory R, Parker T, Cunningham T, Legg C, Lumb A, Parker K, Prud’homme D, Gaboury I, Adamo KB: Влияние открытой обратной связи на физическую активность и просмотр телепередач при избыточном весе и дети с ожирением: рандомизированное контролируемое исследование. Педиатрия. 2006, 118 (1): e157-e166. 10.1542/пед.2005-3052.
Артикул Google ученый
Мэлони А.Э., Бетеа Т.С., Келси К.С., Маркс Дж.Т., Паез С., Розенберг А.М., Кателье Д.Дж., Хамер Р.М., Сикич Л.: Пилотная версия видеоигры (DDR) для поощрения физической активности и сокращения сидячего времени перед экраном. Ожирение. 2008, 16 (9)): 2074-2080. 10.1038/обык.2008.295.
Артикул Google ученый
Ни Мхурчу С., Робертс В., Мэддисон Р., Дори Э., Цзян И., Джулл А., Тин Тин С. Влияние электронных часов на просмотр телевизора детьми: экспериментальное испытание вмешательства на дому. Пред. мед. 2009, 49 (5): 413-417. 10.1016/j.ypmed.2009.
09.003.Артикул Google ученый
Рёммих Дж. Н., Гургол С. М., Эпштейн Л. Х.: Обратная связь без обратной связи повышает физическую активность молодежи. Медицинские спортивные упражнения. 2004, 36 (4): 668-673. 10.1249/01.МСС.0000121947.59529.3Б.
Артикул Google ученый
Maddison R, Mhurchu CN, Foley L, Epstein L, Jiang Y, Tsai M, Dewes O, Heke I: Экранное вмешательство по снижению веса, ориентированное на детей дома (SWITCH): протокол рандомизированного контролируемого исследования. Здоровье в пабе BMC. 2011, 11: 524-10.1186/1471-2458-11-524.
Артикул Google ученый
Cole TJ, Flegal KM, Nicholls D, Jackson AA: Предельные значения индекса массы тела для определения худобы у детей и подростков: международное исследование. БМЖ. 2007, 335 (7612): 194-10.
1136/bmj.39238.399444.55.Артикул Google ученый
Исследование состояния здоровья Новой Зеландии: ежегодное обновление основных результатов за 2012–2013 гг. 2013, Веллингтон, Министерство здравоохранения
Бандура А, Бандура А: Социальные основы мысли и действия: когнитивная социальная теория. 1986, Pretince Hall, Englewood Cliffs, New York
Google ученый
Хурш С.Р., Бауман Р.А.: Поведенческий анализ спроса. Adv Behav Econ. 1987, 1: 117-165.
Google ученый
Эпштейн Л.Х., Рёммих Ю.Н.: Снижение малоподвижного образа жизни: роль в изменении физической активности. Exerc Sport Sci Rev. 2001, 29 (3): 103-108. 10.1097/00003677-200107000-00003.
КАС Статья Google ученый
Эпштейн Л.Х., Саленс Б.Е., Майерс М.Д., Вито Д.: Влияние снижения малоподвижного образа жизни на выбор деятельности у детей с ожирением. Психология здоровья. 1997, 16 (2): 107-113. 10.1037/0278-6133.16.2.107.
КАС Статья Google ученый
Saelens BE, Epstein LH: Поведенческая инженерия выбора деятельности у детей с ожирением. Int J Obes Relat Metab Disord. 1998, 22 (3): 275-277. 10.1038/sj.ijo.0800570.
КАС Статья Google ученый
Эпштейн Л.Х., Палух Р.А., Горди К.С., Дорн Дж.
: Снижение малоподвижного образа жизни при лечении детского ожирения. Arch Pediatr Adolesc Med. 2000, 154 (3): 220-226. 10.1001/архпеди.154.3.220.КАС Статья Google ученый
Нортон К., Олдс Т.: Антропометрика. 1996, Университет Нового Южного Уэльса, Сидней, Австралия
Google ученый
Раш Э., Пуниани К., Валенсия М.Э., Дэвис П.С.В., Планк Л.Д.: Оценка упитанности тела по индексу массы тела и биоэлектрическому импедансу: сравнение новозеландских детей из Европы, маори и детей с островов Тихого океана. Eur J Clin Nutr. 2003, 57: 1394-1401. 10.1038/sj.ejcn.1601701.
КАС Статья Google ученый
Ридли К., Олдс Т.С., Хилл А., Ридли К., Олдс Т.С., Хилл А.: Воспоминание о мультимедийных занятиях для детей и подростков (MARCA): разработка и оценка.
Int J Behav Nutr Phys Act. 2006, 3 (1): 10-10.1186/1479-5868-3-10.Артикул Google ученый
Бут М.Л., Эйнсворт Б.Е., Пратт М., Экелунд У., Ингве А., Саллис Дж.Ф., Оджа П.: Международный опросник по физической активности: надежность и валидность в 12 странах. Медицинские спортивные упражнения. 2003, 195 (9131/03): 3508.
Google ученый
NZ Food, NZ Children. Основные результаты Национального исследования питания детей 2002 года. 2003 г., Министерство здравоохранения, Веллингтон, Новая Зеландия
Меткалф П., Скрэгг Р., Шарп С., Фитцджеральд Э., Шааф Д., Уоттс К.: Кратковременная повторяемость опросника частоты приема пищи в Новой Зеландии Дети в возрасте 1–14 лет. Eur J Clin Nutr. 2003, 57: 1498-1503. 10.1038/sj.ejcn.1601717.
КАС Статья Google ученый
Motl RW, Dishman RK, Saunders R, Dowda M, Felton G, Pate RR: Измерение удовольствия от физической активности у девочек-подростков. Am J Prev Med. 2001, 21 (2): 110-117. 10.1016/S0749-3797(01)00326-9.
КАС Статья Google ученый
Салмон Дж., Оуэн Н., Кроуфорд Д., Бауман А., Саллис Дж. Ф.: Физическая активность и сидячий образ жизни: популяционное исследование барьеров, удовольствия и предпочтений. Психология здоровья. 2003, 22 (2): 178-188. 10.1037/0278-6133.22.2.178.
Артикул Google ученый
Марш С., Фоули Л.С., Уилкс Д.К., Мэддисон Р.: Семейные вмешательства для сокращения малоподвижного образа жизни у молодежи: систематический обзор рандомизированных контролируемых испытаний.
Obes Rev. 2013, 15 (2): 117-133. 10.1111/обр.12105.Артикул Google ученый
Робисон Д.Л.: Вознаграждать?… или не вознаграждать?: подвергая сомнению целесообразность использования внешнего подкрепления в программах укрепления здоровья. Am J Health Promot. 1998, 13 (1): 1-3. 10.4278/0890-1171-13.1.1.
КАС Статья Google ученый
Тимперио А., Салмон Дж., Болл К.: Основанные на фактических данных стратегии поощрения физической активности среди детей, подростков и молодых людей: обзор и обновление. J Sci Med Sport. 2004, 7 (1 Приложение): 20-29. 10.1016/С1440-2440(04)80274-3.
КАС Статья Google ученый
Zwarenstein M, Treweek S, Gagnier JJ, Altman DG, Tunis S, Haynes B, Oxman AD, Moher D: Улучшение отчетности о практических испытаниях: расширение заявления CONSORT.
БМЖ. 2008, 337: а2390-10.1136/bmj.a2390.Артикул Google ученый
Sonneville KR, Rifas-Shiman SL, Kleinman KP, Gortmaker SL, Gillman MW, Taveras EM: Ассоциации поведения, вызывающего ожирение, у матерей и детей с ожирением, участвующих в рандомизированном исследовании. Ожирение. 2012, 20 (7): 1449-1454. 10.1038/об.2012.43.
Артикул Google ученый
Робертс Д.Ф., Фёр Ю.Г.: Тенденции использования медиа. Будущее дитя. 2008, 18 (1): 11-37. 10.1353/фок.0.0000.
Артикул Google ученый
Национальный институт инноваций в области здравоохранения Оклендского университета, Private Bag , Окленд, 1142, Новая Зеландия
Ральф Мэддисон, Саманта Марш, Карен Картер, Яннан Цзян и Клиона Ни Мхурчу
Центр Великобритании и CRC 900 Исследование деятельности, Школа клинической медицины Кембриджского университета, Кембридж, АнглияКафедры педиатрии, общественного здравоохранения и поведения в отношении здоровья, социальной и профилактической медицины, Школа медицины и биомедицинских наук Университета Буффало, Буффало, США
Leonard H Epstein
School of Health Sciences, University of South Australia, Adelaide, Australia
Timothy Olds
Pacific Health, University of Auckland, Auckland, New Zealand
Ofa Dewes
Heke Consulting, Окленд, Новая Зеландия
Ихранги Хеке
- Ральф Мэддисон
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Samantha Marsh
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Louise Foley
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Leonard H Epstein
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Тимоти Олдс
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ofa Dewes
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ihirangi Heke
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Карен Картер
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yannan Jiang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Cliona Ni Mhurchu
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
CrossRef Full Text | Google Scholar
76. Baratelli F, Lin Y, Zhu L, Yang SC, Heuze-Vourc’h N, Zeng G, et al. Простагландин E2 индуцирует экспрессию гена FOXP3 и функцию Т-регуляторных клеток в Т-клетках CD4+ человека. Дж Иммунол. (2005) 175:1483–90. doi: 10.4049/jimmunol.175.3.1483
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
77. Pan PY, Ma G, Weber KJ, Ozao-Choy J, Wang G, Yin B, et al. Иммуностимулирующий рецептор CD40 необходим для подавления Т-клеток и активации Т-регуляторных клеток, опосредованной супрессорными клетками миелоидного происхождения при раке. Рак Res . (2010) 70:99–108. doi: 10. 1158/0008-5472.CAN-09-1882
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
78. Zoso A, Mazza EM, Bicciato S, Mandruzzato S, Bronte V, Serafini P, et al. Супрессорные клетки человеческого фиброцитарного миелоидного происхождения экспрессируют IDO и способствуют толерантности за счет экспансии Treg-клеток. Евро J Иммунол . (2014) 44:3307–19. doi: 10.1002/eji.201444522
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
79. Ерусланов Э.Б., Бходжнагарвала П.С., Кватромони Дж.Г., Стивен Т.Л., Ранганатан А., Дешпанде С. и соавт. Ассоциированные с опухолью нейтрофилы стимулируют Т-клеточный ответ при раке легкого человека на ранней стадии. Дж Клин Инвест. (2014) 124:5466–80. doi: 10.1172/JCI77053
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
80. Mancino A, Schioppa T, Larghi P, Pasqualini F, Nebuloni M, Chen IH, et al. Различные эффекты гипоксии на функции дендритных клеток. Кровь (2008) 112:3723–34. дои: 10.1182/кровь-2008-02-142091
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
81. Элиа А.Р., Каппелло П., Пуппо М., Фраоне Т., Ванни С., Ева А. и др. Дендритные клетки человека, дифференцированные в условиях гипоксии, подавляют поглощение антигена и изменяют свой профиль экспрессии хемокинов. J Лейкок Биол. (2008) 84:1472–82. doi: 10.1189/jlb.0208082
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
82. Yang M, Ma C, Liu S, Shao Q, Gao W, Song B и др. HIF-зависимая индукция аденозинового рецептора A2b искажает дендритные клетки человека до Th3-стимулирующего фенотипа при гипоксии. Иммунол Селл Биол . (2010) 88:165–71. doi: 10.1038/icb.2009.77
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
83. Ghiringhelli F, Puig PE, Roux S, Parcellier A, Schmitt E, Solary E, et al. Опухолевые клетки превращают незрелые миелоидные дендритные клетки в TGF-бета-секретирующие клетки, индуцирующие пролиферацию регуляторных CD4+CD25+ Т-клеток. J Exp Med. (2005) 202:919–29. doi: 10.1084/jem.20050463
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
84. Munn DH, Sharma MD, Lee JR, Jhaver KG, Johnson TS, Keskin DB, et al. Потенциальная регуляторная функция дендритных клеток человека, экспрессирующих индоламин-2,3-диоксигеназу. Наука (2002) 297:1867–70. doi: 10.1126/science.1073514
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
85. Меллор А.Л., Манн Д.Х. Экспрессия IDO дендритными клетками: толерантность и катаболизм триптофана. Nat Rev Immunol. (2004) 4:762–74. doi: 10.1038/nri1457
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
86. Новицкий С.В., Рыжов С., Зайнагетдинов Р., Гольдштейн А.Е., Хуанг Ю., Тихомиров О.Ю. Аденозиновые рецепторы в регуляции дифференцировки и функции дендритных клеток. Кровь (2008) 112:1822–31. doi: 10.1182/blood-2008-02-136325
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
87. Liu Q, Zhang C, Sun A, Zheng Y, Wang L, Cao X. Регуляторные дендритные клетки CD11bhighIalow, образованные опухолью, подавляют Т-клеточный ответ посредством аргиназы I. J Иммунол. (2009) 182:6207–16. doi: 10.4049/jimmunol.0803926
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
88. Альбини А., Бруно А., Нунан Д.М., Мортара Л. Вклад в ангиогенез опухоли клеток врожденного иммунитета в микроокружении опухоли: последствия для иммунотерапии. Фронт Иммунол. (2018) 9:527. doi: 10.3389/fimmu.2018.00527
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
89. Гордон-Уикс А.Н., Лим С.Ю., Южалин А.Е., Джонс К., Маркелк Б., Ким К.Дж. и соавт. Нейтрофилы способствуют росту метастазов в печени посредством зависимого от фактора роста 2 фибробластов ангиогенеза у мышей. Гепатология (2017) 65:1920–35. doi: 10.1002/hep.29088
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
90. Tabaries S, Ouellet V, Hsu BE, Annis MG, Rose AA, Meunier L, et al. Гранулоцитарные иммунные инфильтраты необходимы для эффективного образования метастазов рака молочной железы в печень. Рак молочной железы Res . (2015) 17:45. doi: 10.1186/s13058-015-0558-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
91. Kowanetz M, Wu X, Lee J, Tan M, Hagenbeek T, Qu X, et al. Гранулоцитарно-колониестимулирующий фактор способствует метастазированию в легкие за счет мобилизации гранулоцитов Ly6G+Ly6C+. Proc Natl Acad Sci USA. (2010) 107:21248–55. doi: 10.1073/pnas.1015855107
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
92. Shi H, Zhang J, Han X, Li H, Xie M, Sun Y и др. Рекрутированные моноцитарные супрессорные клетки миелоидного происхождения способствуют аресту опухолевых клеток в преметастатической нише за счет опосредованного ИЛ-1бета увеличения экспрессии Е-селектина. Int J Рак . (2017) 140:1370–83. doi: 10.1002/ijc.30538
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
93. Safarzadeh E, Orangi M, Mohammadi H, Babaie F, Baradaran B. Супрессорные клетки миелоидного происхождения: важные факторы, способствующие прогрессированию опухоли и метастазированию. J Cell Physiol. (2018) 233:3024–36. doi: 10.1002/jcp.26075
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
94. Golden EB, Frances D, Pellicciotta I, Demaria S, Helen Barcellos-Hoff M, Formenti SC. Радиация способствует дозозависимой и индуцированной химиотерапией иммуногенной гибели клеток. Онкоиммунология (2014) 3:e28518. doi: 10.4161/onci.28518
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
95. Burnette BC, Liang H, Lee Y, Chlewicki L, Ходарев NN, Weichselbaum RR, et al. Эффективность лучевой терапии зависит от индукции интерферон-зависимого врожденного и адаптивного иммунитета I типа. Рак Res . (2011) 71:2488–96. doi: 10.1158/0008-5472.CAN-10-2820
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
96. Гупта А., Пробст Х.К., Вуонг В., Ландшаммер А., Мут С., Ягита Х. и др. Лучевая терапия способствует развитию опухолеспецифических эффекторных CD8+ Т-клеток посредством активации дендритных клеток. Дж Иммунол . (2012) 189: 558–66. doi: 10.4049/jimmunol.1200563
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
97. Klug F, Prakash H, Huber PE, Seibel T, Bender N, Halama N, et al. Облучение в малых дозах программирует дифференцировку макрофагов до фенотипа iNOS(+)/M1, что обеспечивает эффективную иммунотерапию Т-клетками. Раковая клетка. (2013) 24:589–602. doi: 10.1016/j.ccr.2013.09.014
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
98. Formenti SC, Demaria S. Объединение лучевой терапии и иммунотерапии рака: сдвиг парадигмы. J Natl Cancer Inst. (2013) 105: 256–65. doi: 10.1093/jnci/djs629
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
99. Рассел Дж.С., Браун Дж.М. Микроокружение облученной опухоли: роль опухолеассоциированных макрофагов в восстановлении сосудов. Фронт Физиол. (2013) 4:157. doi: 10.3389/fphys.2013.00157
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
100. Ватнер Р.Э., Форменти СК. Миелоидные клетки в опухолях: эффекты радиации. Semin Radiat Oncol . (2015) 25:18–27. doi: 10.1016/j.semradonc.2014.07.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
101. Xu J, Escamilla J, Mok S, David J, Priceman S, West B, et al. Блокада передачи сигналов CSF1R останавливает инфильтрирующие опухоль миелоидные клетки и повышает эффективность лучевой терапии при раке предстательной железы. Рак Res. (2013) 73: 2782–94. doi: 10.1158/0008-5472.CAN-12-3981
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
102. Шиао С.Л., Раффелл Б., ДеНардо Д.Г., Фаддегон Б.А., Парк К.С., Куссенс Л.М. Th3-поляризованные CD4(+) Т-клетки и макрофаги ограничивают эффективность лучевой терапии. Рак Иммунол Рез . (2015) 3: 518–25. doi: 10.1158/2326-6066.CIR-14-0232
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
103. Deng L, Liang H, Burnette B, Beckett M, Darga T, Weichselbaum RR, et al. Облучение и лечение анти-PD-L1 синергически повышают противоопухолевый иммунитет у мышей. Дж Клин Инвест. (2014) 124:687–95. doi: 10.1172/JCI67313
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
104. Ma Y, Galluzzi L, Zitvogel L, Kroemer G. Аутофагия и клеточные иммунные реакции. Иммунитет (2013) 39:211–27. doi: 10.1016/j.immuni.2013.07.017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
105. Пфиршке С., Энгблом С., Рикельт С., Кортез-Ретамозо В., Гаррис С., Пуччи Ф. и др. Иммуногенная химиотерапия делает опухоли чувствительными к терапии блокадой контрольных точек. Иммунитет (2016) 44:343–54. doi: 10.1016/j.immuni.2015.11.024
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
106. Miller MA, Zheng YR, Gadde S, Pfirschke C, Zope H, Engblom C, et al. Ассоциированные с опухолью макрофаги действуют как медленно высвобождаемый резервуар нанотерапевтического пролекарства Pt(IV). Нац.коммун. (2015) 6:8692. doi: 10.1038/ncomms9692
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
107. Иида Н., Дзуцев А., Стюарт К.А., Смит Л., Буладу Н., Вайнгартен Р.А. и соавт. Комменсальные бактерии контролируют реакцию рака на терапию, модулируя микроокружение опухоли. Наука (2013) 342:967–70. doi: 10.1126/science.1240527
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
108. Vincent J, Mignot G, Chalmin F, Ladoire S, Brucard M, Chevriaux A, et al. 5-фторурацил избирательно убивает ассоциированные с опухолью супрессорные клетки миелоидного происхождения, что приводит к усилению Т-клеточно-зависимого противоопухолевого иммунитета. Рак Res. (2010) 70:3052–61. doi: 10.1158/0008-5472.CAN-09-3690
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
109. DeNardo DG, Brennan DJ, Rexhepaj E, Ruffell B, Shiao SL, Madden SF, et al. Сложность лейкоцитов предсказывает выживаемость рака молочной железы и функционально регулирует ответ на химиотерапию. Рак Дисков . (2011) 1:54–67. doi: 10.1158/2159-8274.CD-10-0028
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
110. Bonapace L, Coissieux MM, Wyckoff J, Mertz KD, Varga Z, Junt T, et al. Прекращение ингибирования CCL2 ускоряет метастазирование рака молочной железы, способствуя ангиогенезу. Природа (2014) 515:130–3. doi: 10.1038/nature13862
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
111. Shree T, Olson OC, Elie BT, Kester JC, Garfall AL, Simpson K, et al. Макрофаги и катепсиновые протеазы притупляют химиотерапевтический ответ при раке молочной железы. Гены Дев . (2011) 25:2465–79. doi: 10.1101/gad.180331.111
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
112. Брушар М., Миньо Г., Деранжер В., Чалмин Ф., Шеврио А., Вегран Ф. и др. Запускаемое химиотерапией высвобождение катепсина В в супрессорных клетках миелоидного происхождения активирует инфламмасому Nlrp3 и способствует росту опухоли. Nat Med . (2013) 19:57–64. doi: 10.1038/nm.2999
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
113. Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CM, Pryer N, et al. Макрофаг IL-10 блокирует CD8+ Т-клеточно-зависимые ответы на химиотерапию путем подавления экспрессии IL-12 во внутриопухолевых дендритных клетках. Раковая клетка (2014) 26:623–37. doi: 10.1016/j.ccell.2014.09.006
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
114. Mo HN, Liu P. Ориентация на МЕТ в терапии рака. Хронический Dis Transl Med . (2017) 3:148–53. doi: 10.1016/j.cdtm.2017.06.002
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
115. Neuzillet C, Tijeras-Raballand A, Cohen R, Cros J, Faivre S, Raymond E, et al. Ориентация на путь TGFbeta для лечения рака. Фармакол Тер . (2015) 147:22–31. doi: 10.1016/j.pharmthera.2014.11.001
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
116. Cavnar MJ, Zeng S, Kim TS, Sorenson EC, Ocuin LM, Balachandran VP, et al. Ингибирование онкогена KIT вызывает поляризацию внутриопухолевых макрофагов M2. J Exp Med. (2013) 210:2873–86. doi: 10.1084/jem.20130875
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
117. Hoeflich KP, O’Brien C, Boyd Z, Cavet G, Guerrero S, Jung K, et al. Противоопухолевая активность MEK и ингибиторов фосфатидилинозитол-3-киназы in vivo в моделях базальноподобного рака молочной железы. Клин Рак Рез . (2009) 15:4649–64. doi: 10. 1158/1078-0432.CCR-09-0317
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
118. McCubrey JA, Steelman LS, Chappell WH, Abrams SL, Wong EW, Chang F, et al. Роль пути Raf/MEK/ERK в росте клеток, злокачественной трансформации и лекарственной устойчивости. Biochim Biophys Acta (2007) 1773:1263–84. doi: 10.1016/j.bbamcr.2006.10.001
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
119. Ван П.Т., Гарнетт М.Дж., Роу С.М., Ли С., Никулеску-Дуваз Д., Гуд В.М. и соавт. Механизм активации сигнального пути RAF-ERK онкогенными мутациями B-RAF. Cell (2004) 116:855–67. doi: 10.1016/S0092-8674(04)00215-6
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
120. Чжан Б.Х., Гуань К.Л. Для активации киназы B-Raf требуется фосфорилирование консервативных остатков Thr598 и Ser601. Эмбо Дж. (2000) 19:5429–39. дои: 10.1093/emboj/19.20.5429
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
121. Steinberg SM, Shabaneh TB, Zhang P, Martyanov V, Li Z, Malik BT, et al. Миелоидные клетки, нарушающие иммунотерапию, восстанавливаются в меланомах с приобретенной резистентностью к ингибиторам BRAF. Рак Res. (2017) 77:1599–610. doi: 10.1158/0008-5472.CAN-16-1755
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
122. Smith MP, Sanchez-Laorden B, O’Brien K, Brunton H, Ferguson J, Young H, et al. Иммунное микроокружение придает устойчивость к ингибиторам пути MAPK через TNF-альфа, полученный из макрофагов. Рак Дисков . (2014) 4:1214–29. doi: 10.1158/2159-8290.CD-13-1007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
123. Mok S, Tsoi J, Koya RC, Hu-Lieskovan S, West BL, Bollag G, et al. Ингибирование рецептора колониестимулирующего фактора-1 повышает противоопухолевую эффективность ингибирования BRAF. BMC Рак (2015) 15:356. doi: 10.1186/s12885-015-1377-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
124. Ott PA, Henry T, Baranda SJ, Frleta D, Manches O, Bogunovic D, et al. Ингибирование как BRAF, так и MEK при меланоме с мутацией BRAF(V600E) восстанавливает нарушенную функцию дендритных клеток (ДК), оказывая при этом различные прямые эффекты на свойства ДК. Рак Иммунол Иммунотер . (2013) 62:811–22. doi: 10.1007/s00262-012-1389-z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
125. Ott PA, Bhardwaj N. Влияние активации пути MAPK при меланоме BRAF(V600) на функцию Т-клеток и дендритных клеток. Фронт Иммунол. (2013) 4:346. doi: 10.3389/fimmu.2013.00346
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
126. Finisguerra V, Di Conza G, Di Matteo M, Serneels J, Costa S, Thompson AA, et al. МЕТ необходим для рекрутирования противоопухолевых нейтрофилов. Природа (2015) 522:349–53. doi: 10.1038/nature14407
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
127. Пардолл Д. М. Блокада иммунных контрольных точек при иммунотерапии рака. Nat Rev Cancer (2012) 12:252–64. doi: 10.1038/nrc3239
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
128. Топалян С.Л., Дрейк К.Г., Пардолл Д.М. Блокада иммунных контрольных точек: общий подход к терапии рака. Раковая клетка (2015) 27:450–61. doi: 10.1016/j.ccell.2015.03.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
129. Шарма П., Ху-Лиескован С., Варго Дж. А., Рибас А. Первичная, адаптивная и приобретенная устойчивость к иммунотерапии рака. Cell (2017) 168:707–23. doi: 10.1016/j.cell.2017.01.017
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
130. Дженкинс Р.В., Барби Д.А., Флаэрти К.Т. Механизмы резистентности к ингибиторам иммунных контрольных точек. BrJ Рак (2018) 118:9–16. doi: 10.1038/bjc.2017.434
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
131. Meyer C, Cagnon L, Costa-Nunes CM, Baumgaertner P, Montandon N, Leyvraz L, et al. Частота циркулирующих MDSC коррелирует с клиническим исходом у пациентов с меланомой, получавших ипилимумаб. Рак Иммунол Иммунотер . (2014) 63:247–57. doi: 10.1007/s00262-013-1508-5
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
132. Бьорн Дж., Юул Ничке Н., Зееберг Иверсен Т., Шмидт Х., Фоде К., Сване ИМ. Иммунологические корреляты лечения и ответа у пациентов со злокачественной меланомой IV стадии, получавших ипилимумаб. Онкоиммунология (2016) 5:e1100788. doi: 10.1080/2162402X.2015.1100788
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
133. Highfill SL, Cui Y, Giles AJ, Smith JP, Zhang H, Morse E, et al. Нарушение CXCR2-опосредованного переноса опухоли MDSC повышает эффективность анти-PD1. Sci Transl Med. (2014) 6:237ra67. doi: 10.1126/scitranslmed.3007974
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
134. Orillion A, Hashimoto A, Damayanti NP, Shen L, Adelaiye-Ogala R, Arisa S, et al. Энтиностат нейтрализует супрессорные клетки миелоидного происхождения и усиливает противоопухолевый эффект ингибирования PD-1 в мышиных моделях карциномы легкого и почки. Клин Рак Res . (2017) 23:5187–201. doi: 10.1158/1078-0432.CCR-17-0741
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
135. Kim K, Skora AD, Li Z, Liu Q, Tam AJ, Blosser RL, et al. Эрадикация метастатического рака у мышей, устойчивого к блокаде иммунных контрольных точек, путем подавления клеток миелоидного происхождения. Proc Natl Acad Sci. (2014) 111:11774. doi: 10.1073/pnas.1410626111
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
136. Arlauckas SP, Garris CS, Kohler RH, Kitaoka M, Cuccarese MF, Yang KS, et al. Визуализация in vivo выявляет ассоциированный с опухолью путь резистентности, опосредованный макрофагами, при терапии анти-PD-1. Sci Trans Med . (2017) 9: eaal3604. doi: 10.1126/scitranslmed.aal3604
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
137. Noman MZ, Desantis G, Janji B, Hasmim M, Karray S, Dessen P, et al. PD-L1 является новой прямой мишенью HIF-1альфа, и его блокада при гипоксии усиливает активацию Т-клеток, опосредованную MDSC. J Exp Med. (2014) 211: 781–90. doi: 10.1084/jem.20131916
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
138. Porta C, Sica A, Riboldi E. Опухолеассоциированные миелоидные клетки: новое понимание их метаболической регуляции и их влияние на иммунотерапию рака. Febs J. (2018) 285:717–33. doi: 10.1111/febs.14288
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
139. Kuang DM, Zhao Q, Peng C, Xu J, Zhang JP, Wu C, et al. Активированные моноциты в перитуморальной строме гепатоцеллюлярной карциномы способствуют иммунной привилегии и прогрессированию заболевания через PD-L1. J Exp Med. (2009) 206:1327–37. doi: 10.1084/jem.20082173
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
140. Curiel TJ, Wei S, Dong H, Alvarez X, Cheng P, Mottram P, et al. Блокада B7-h2 улучшает противоопухолевый иммунитет, опосредованный миелоидными дендритными клетками. Нац. мед. (2003) 9: 562–7. doi: 10.1038/nm863
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
141. Spranger S, Bao R, Gajewski TF. Присущая меланоме передача сигналов бета-катенина предотвращает противоопухолевый иммунитет. Природа (2015) 523:231–5. doi: 10.1038/nature14404
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
142. Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau CS, et al. ИЛ-17-продуцирующие гамма-дельта Т-клетки и нейтрофилы способствуют метастазированию рака молочной железы. Природа (2015) 522:345–8. doi: 10.1038/nature14282
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
143. Шривастава М.К., Чжу Л., Харрис-Уайт М., Кар Великобритания, Хуанг М., Джонсон М.Ф. и др. Истощение миелоидных супрессорных клеток увеличивает противоопухолевую активность при раке легкого. PLoS ONE (2012) 7:e40677. doi: 10.1371/annotation/5c756e7d-6e97-416f-836a-dced97cf46af
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
144. Qian BZ, Li J, Zhang H, Kitamura T, Zhang J, Campion LR, et al. CCL2 рекрутирует воспалительные моноциты, чтобы облегчить метастазирование опухоли молочной железы. Природа (2011) 475:222–5. doi: 10.1038/nature10138
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
145. Zhu Y, Knolhoff BL, Meyer MA, Nywening TM, West BL, Luo J, et al. Блокада CSF1/CSF1R перепрограммирует инфильтрирующие опухоль макрофаги и улучшает ответ на иммунотерапию Т-клеточными контрольными точками в моделях рака поджелудочной железы. Рак Res. (2014) 74:5057–69. doi: 10.1158/0008-5472.CAN-13-3723
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
146. Ries CH, Cannarile MA, Hoves S, Benz J, Wartha K, Runza V, et al. Нацеливание на ассоциированные с опухолью макрофаги с помощью антител против CSF-1R раскрывает стратегию терапии рака. Раковая клетка (2014) 25:846–59. doi: 10.1016/j.ccr.2014.05.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
147. Strachan DC, Ruffell B, Oei Y, Bissell MJ, Coussens LM, Pryer N, et al. Ингибирование CSF1R задерживает рост опухоли шейки матки и молочной железы в мышиных моделях за счет ослабления оборота ассоциированных с опухолью макрофагов и усиления инфильтрации CD8(+) Т-клетками. Онкоиммунология (2013) 2:e26968. doi: 10.4161/onci.26968
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
148. Mitchem JB, Brennan DJ, Knolhoff BL, Belt BA, Zhu Y, Sanford DE, et al. Нацеливание на макрофаги, инфильтрирующие опухоль, уменьшает количество инициирующих опухоль клеток, снимает иммуносупрессию и улучшает химиотерапевтические реакции. Рак Res . (2013) 73:1128–41. doi: 10.1158/0008-5472.CAN-12-2731
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
149. Pyonteck SM, Akkari L, Schuhmacher AJ, Bowman RL, Sevenich L, Quail DF, et al. Ингибирование CSF-1R изменяет поляризацию макрофагов и блокирует прогрессирование глиомы. Нац. мед. (2013) 19:1264–72. doi: 10.1038/nm.3337
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
150. Chen X, Wang Y, Nelson D, Tian S, Mulvey E, Patel B, et al. CCL2/CCR2 регулирует микроокружение опухоли в карциномах молочной железы, вызванных HER-2/neu, у мышей. PLoS ONE (2016) 11:e0165595. doi: 10.1371/journal.pone.0165595
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
151. Лесохин А.М., Холь Т.М., Китано С., Кортез С., Хиршхорн-Симерман Д., Авогадри Ф. и др. Моноцитарные супрессорные клетки CCR2(+) миелоидного происхождения способствуют ускользанию от иммунного ответа, ограничивая инфильтрацию активированных CD8 Т-клеток в микроокружение опухоли. Рак Res . (2012) 72:876–86. doi: 10.1158/0008-5472.CAN-11-1792
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
152. Izhak L, Wildbaum G, Weinberg U, Shaked Y, Alami J, Dumont D, et al. Преобладающая экспрессия CCL2 в месте опухоли у пациентов с раком предстательной железы приводит к селективной потере иммунологической толерантности к CCL2, которая может быть амплифицирована полезным образом. Дж Иммунол . (2010) 184:1092–101. doi: 10.4049/jimmunol.10
Нельсон М.С., Ноймарк-Штайнер Д., Ханнан П.Дж., Сирард Дж.Р., Стори М.: Продольные и долгосрочные тенденции физической активности и малоподвижного образа жизни в подростковом возрасте. Педиатрия. 2006, 118 (6): e1627-e1634. 10.1542/пед.2006-0926.
Артикул Google ученый
Maddison R, Foley L, Mhurchu CN, Jiang Y, Jull A, Prapavessis H, Hohepa M, Rodgers A: Влияние активных видеоигр на состав тела: рандомизированное контролируемое исследование. Am J Clin Nutr. 2011, 94 (1): 156-163. 10.3945/ajcn.110.009142.
КАС Статья Google ученый
Эпштейн Л.Х., Саленс Б.Е., О’Брайен Дж.Г.: Эффекты усиления активного поведения по сравнению с уменьшением малоподвижного поведения у детей с ожирением. Int J Behav Med. 1995, 2 (1): 41-50. 10.1207/s15327558ijbm0201_4.
КАС Статья Google ученый
Kendzierski D, DeCarlo KJ: Шкала удовольствия от физической активности: Два проверочных исследования. J Sport Exerc Psychol. 1991, 13 (1): 50-64.
Google ученый
Загрузить ссылки
Благодарности
Это было исследование, инициированное исследователем, финансируемое за счет гранта Совета по исследованиям в области здравоохранения Новой Зеландии (10/). Докторская стипендия Bright Futures Комиссии по высшему образованию. CNM была поддержана стипендией Heart Foundation of New Zealand Senior Fellowship (грант 1380). Авторы хотели бы поблагодарить всех, кто участвовал в этом испытании.
Источники поддержки
Это исследование финансировалось Советом по исследованиям в области здравоохранения Новой Зеландии (10/077). Д-р Ральф Мэддисон получил поддержку исследовательской стипендии Heart Foundation (грант 1211). Профессор Клиона Ни Мхурчу получила поддержку старшего товарищества Национального фонда сердца (грант 1380). Д-р Луиза Фоли получила поддержку от Фонда Сердца Новой Зеландии.
Информация об авторе
Авторы и организации
Louise Foley
Авторы
Автор, ответственный за переписку
Соответствие
Ральф Мэддисон.