
Какой размер обложки в ВК 2022?
Визуальное оформление личного профиля или сообщества в VK – важная составляющая их успеха, роста посещаемости и дальнейшего продвижения. Запуск обложки для личного профиля в ВК в октябре 2022 года, различные изменения требований к обложке в группах, появление живых обложек требуют знания точных размеров для обложек в ВК. Какие должны быть размеру обложек (шапок) в социальной сети «ВКонтакте» в 2022 году мы и расскажем в нашем материале.
Содержание
- Размеры для обложки личного профиля в ВК
- Размеры для шапки сообщества в VK
- Размеры для живой обложки в ВКонтакте
- Видео
Размеры для обложки личного профиля в ВК
Как известно, 10 октября 2022 года социальная сеть «ВКонтакте» в честь своего 16-летия подарила своим пользователям возможность ставить обложку на свои личные страницы в ВК. Раньше такая возможность присутствовала лишь у сообществ и публичных страниц, но теперь любой пользователь VK получил возможность украсить свой аккаунт яркой и привлекающей внимания обложкой.
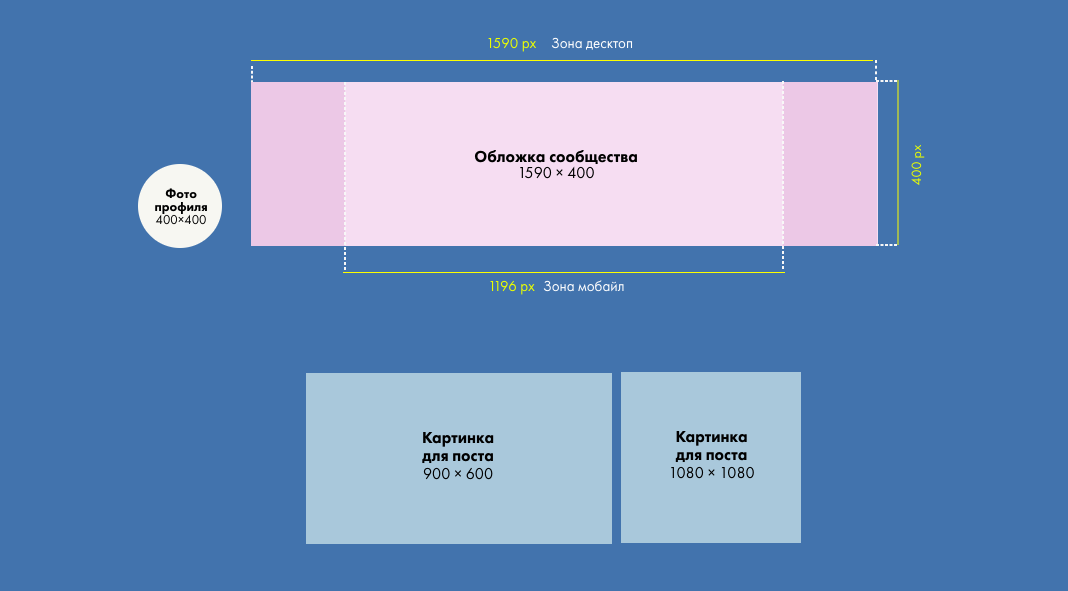
Оптимальное разрешение для обложки личного профиля в ВК — 1920×640 пикселей, формат файла – JPG, PNG или WEBP.
Для загрузки обложки для личного профиля в ВК необходимо перейти на главную страницу своего профиля, нажмите справа сверху на «Изменить обложку» — «Загрузить изображение».
При этом функционал ВК позволяет выбрать область обложки, если визуальная часть загруженного изображения выходит за рекомендуемые границы.
Читайте также: обложки для групп ВК 1590×400.
Размеры для шапки сообщества в VK
В случае сообществ (групп) в ВК оптимальный размер обложки составляет 1590 на 530 пикселей. В этом случае шапка будет хорошо отображаться на экране ПК. В случае загрузки большего по размеру изображения ВКонтакте позволяет автоматически выделить нужную область для размещения в форме обложки.
Однако на мобильном устройстве из-за специфики его отображения видимая часть обложки будет составлять 1196×315 пикселей. Из-за такой обрезки, и преобладания мобильных устройство при работе с ВК, рекомендуем размещать важную информацию на шапке вашей группы в границах видимой области для мобильных устройств – 1196 на 315 пикселей.
Для добавления вашей обложки с компьютера выполните следующее:
- Перейдите в вашу группу ВК;
- Выберите там раздел «Управление»;
- В опции «Обложка» нажмите на надпись «Добавить»;
- Загрузите изображение для вашей шапки в ВК нажав на «Добавить обложку».
Для загрузки фото или видео в обложку сообщества на телефоне выполните следующее:
- Перейдите в своё сообщество на телефоне;
- Нажмите на значок шестерёнки в правом верхнем углу;
- Выберите раздел «Информация»;
- Тапните на «Обложка»;
- Добавьте нужную обложку из вашей Галереи нажав на «Загрузить обложку».
Также функционал ВКонтакте позволяет добавить специальную обложку для смартфонов. Размер такой обложки – 1080 на 1920 пикселей.
Размеры для живой обложки в ВКонтакте
Также вы можете сделать живую обложку для вашего сообщества в ВК. Она выглядит как 5 больших видео или фотографий, которые будут переключаться системой автоматически.
Размеры для живых обложек следующие:
- Разрешение изображений или видео – 1080 на 1920 пикселей;
- Продолжительность видео – не больше 30 секунд;
- Размер загружаемых файлов – не более 20 Мегабайт;
- Видеокодек — H.264;
- Аудиокодек — AAC.
Система предполагает следующие формы загрузки живых обложек:
- Загрузка до 5 видеороликов, которые будут меняться автоматически;
- Загрузка до 5 изображений, которые будут меняться автоматически;
- Загрузка до 5 изображений, которые будут меняться, при этом каждое такое изображение будет увеличиваться.
Для установки живой обложки выполните следующее:
- Перейдите в ваше сообщество;
- Кликните на пункт «Управление» в правой части экрана;
- В меню настроек, в блоке основной информации найдите опцию «Обложка», и кликните рядом с ней на «Добавить»;
- В открывшемся меню добавление найдите раздел «Живая обложка сообщества», и нажмите на «Добавить» для загрузки изображений или видео.

Это может пригодится: как сделать исчезающие сообщения в ВК?
Видео
В нашем материале мы выяснили, какой должен быть размер для обложки в личном профиле и группах ВК в 2022 году. С появлением обложки в личных профилях пользователей последние получили возможность ставить обложку по своему вкусу на своей странице. При размещении обложки руководствуйтесь указанными нами параметрами, что позволит выбрать оптимальную обложку под стиль оформления вашего аккаунта.
Опубликовано в рубрике «Интернет»
Обложка для группы ВК — как сделать, размеры в 2023 году
Дата обновления: 10.11.2022 Время чтения: 5 мин.
Лиана Санамян ЭКСПЕРТ
Редактор и автор портала Otzyvmarketing.
Поделиться
Поделитесь статьей с друзьями и коллегами
Известная пословица «Встречают по одежке» применима и к соцсетям – каким бы интересным и полезным ни был ваш контент в Вконтакте, на него вряд ли обратят внимания без правильного оформления сообщества.
Рассказываем, как правильно сделать и оформить обложку для группы ВК в 2022 году.
Тысячи надёжных каналов для продвижения бизнеса в Telegram
Telega.in — платформа нативных интеграций с блогерами в Telegram. Каждый канал в каталоге проверен вручную, а админы готовы к сотрудничеству — никаких долгих переговоров!
- Выбирайте лучшие каналы в открытом каталоге самостоятельно или с помощью менеджера.
- Планируйте кампании эффективно: интеграции гарантированно будут опубликованы в срок и в том виде, в котором их согласует заказчик.
- Экономьте время: отчёты по каждой кампании предоставляются в удобной таблице со всеми необходимыми данными.

ПЕРЕЙТИ НА САЙТ
Как сделать и поставить обложку в группу ВК
Чтобы сделать обложку для сообщества ВК, нужен графический редактор. Можно использовать:
- Онлайн-сервисы (Wilda, Supa, Flyvi.io)
- Встроенные инструменты компьютера или смартфона (например, Paint в Windows)
- Профессиональные программы (Photoshop, GIMP)
Проще всего использовать онлайн-сервисы – они имеют все нужные функции и их не надо устанавливать.
Самый популярный онлайн-сервис для создания обложек для соцсетей – Canva. Правда, сейчас он недоступен для жителей России. Вы можете использовать его с VPN или выбрать аналог – например, Wilda.
Wilda
Чтобы создать обложку, выберите понравившийся шаблон и нажмите «Редактирование».
Откроется окно графического редактора. В верхней части экрана можно изменить размер шаблона, включить сетку или линейку. Слева находятся основные элементы, которые можно разместить на шаблоне – например, блок, картинка, логотип, текст и т.д.
Когда обложка будет готова, просто скачайте ее (JPG или PNG, формат PDF не подойдет для ВК) и установите в группе Вконтакте.
В Wilda можно бесплатно создать или отредактировать обложку. Но скачать ее получится только после оплаты.
Если вам не понравился Wilda, можете использовать любой другой сервис. Например, Supa. В видео ниже видеоинструкция по работе с ним.
Как должна выглядеть хорошая обложка? Все зависит от сферы бизнеса и назначения самой обложки. Например, на ней можно изобразить ваш офис, лучшие работы, контактные данные и т.д. Чаще всего размещают:
- Заголовок и подзаголовок – обычно здесь дублируют название группы, перечисляют основные услуги или товары;
- Логотип – добавляется для узнаваемости бренда;
- Контакты – номер телефона, адрес, ссылки на другие соцсети;
- Фотографии – целевая аудитория, команда, продукты.
Установить обложку в группе ВК можно с компьютера или с телефона. Рассмотрим оба варианта.
С компьютера
Откройте сообщество Вконтакте и в правой части экрана выберите «Управление». На открывшейся странице в разделе «Настройки» («Основная информация») выберите «Обложка» и нажмите «Добавить».
Тут можно добавить:
- Главную обложку
- Живую обложку (о ней будет ниже)
Нажмите «Добавить обложку» и выберите файл на компьютере.
С телефона
Рассмотрим пример для смартфонов на Android. Интерфейс и порядок действий для iPhone не отличается.
Откройте мобильное приложение Вконтакте, перейдите в сообщество и нажмите значок в виде шестеренки в правом верхнем углу. Выберите раздел «Информация» и на открывшейся странице нажмите «Обложка» (вы увидите пометку «Не загружена»). Нажмите «Загрузить» обложку и выберите изображение на телефоне.
Примеры готовых обложек:
Актуальные размеры обложки в 2022
Требования к обложке для группы Вконтакте:
- Размер изображения – 1590х530 пикселей
- Соотношение сторон – 3:1
- Формат – PNG, JPG или GIF (статичный)
Стандартная обложка для сайта и мобильного приложения имеет одинаковый размер. Но учитывайте важный момент – в верхней части экрана смартфона находятся индикаторы (значки сети, заряда и т.д.) и вырез под камеру. Их точное расположение на разных моделях телефонов отличается. Поэтому вообще не располагайте в верхней части обложки важную информацию (номер телефона, ссылки на другие соцсети) – сделайте небольшой отступ в 120-140 пикселей.
То же самое сделайте и для боковых областей – они затемняются и на них располагаются кнопки навигации (стрелка влево, кнопка вызова меню).
Чтобы сделать обложку нужного размера, используйте графические редакторы (о которых мы писали выше). Если у вас есть готовое изображение, но вы боитесь не «угадать» с размерами – ВК поможет с этим. При загрузке обложки можно использовать инструмент, позволяющий повернуть изображение или выбрать область, которая будет отображаться на обложке.
Живая обложка ВК
Живая обложка ВК – это обложка, которая состоит из 5 изображений или видео, которые автоматически сменяются. Иначе говоря, вместо статичного изображения на странице сообщества другие пользователи видят слайд-шоу или короткий видеоролик.
Требования к живой обложке:
- Размер изображений или видео –
- Соотношение сторон – 9:16
- Размер файла – до 20 Мб
- Длительность – до 30 секунд
- Видеокодек – Н.264, аудиокодек – ААС
Если вы хотите сделать живую обложку из изображений, используйте те же самые графические редакторы, о которых мы говорили выше. Если вас интересует видеообложка, вам понадобится видеоредактор. Проще всего использовать мобильные приложения – например, InShot.
Чтобы добавить живую обложку, перейдите в управление сообществом и нажмите «Добавить обложку» (все шаги описаны выше). Но вместо обычной обложки выберите живую.
В живых обложках можно включить эффект приближения – изображения будут показываться в движении. Если на обложке много текста, лучше отключить эту опцию.
Аватар сообщества
Аватар группы ВК – это изображение, которое находится на странице сообщества и отображается рядом с публикациями в ленте новостей. Есть 2 вида аватаров:
- Полный аватар – изображение выводится на странице сообщества
- Миниатюра – изображение выводится в новостях, списках сообществ и комментариях
Если у группы нет обложки, то вместо нее в десктопной версии Вконтакте будет отображаться полный аватар, а в мобильной версии и приложении – миниатюра. Если у группы есть обложка, то везде будет отображаться только миниатюра.
ЧИТАЙТЕ ТАКЖЕ
Как создать группу в ВК: полный гайд
06.10.2022
Требования к аватару:
- Размер изображения – 200х200 пикселей (для миниатюры), 400х400 пикселей (для полного аватара)
- Формат – JPG, GIF или PNG
Создать аватар можно с помощью любого графического редактора.
В первую очередь, вы создаете полный аватар (даже если у вас есть обложка). Для этого выберите квадратную или прямоугольную (вертикальная ориентация) область, которая будет выводиться на странице. После этого вы создаете миниатюру – для этого нужно выбрать маленькую круглую область.
Как сделать меню в группе
Меню – это инструмент для оформления групп ВК, с помощью которого можно структурировать содержимое группы или выделить важную информацию. Например, в меню можно добавить ссылки на статьи, альбомы, товары. Оно будет отображаться везде – в полной версии сайта, мобильной версии и в приложении.
Требования к меню:
- Размер изображения – 376х256 пикселей
- Формат – JPG, GIF или PNG
- Название – до 20 символов
- Количество ссылок – до 7
Чтобы сделать меню в группе, перейдите в управление сообществом (значок в виде шестеренки) и в разделе «Настройки» выберите вкладку «Меню».
Чтобы добавить ссылку на объект внутри Вконтакте, нажмите кнопку «Новая ссылка». Теперь вы сможете:
- Загрузить обложку для пункта меню
- Ввести название
- Указать ссылку
Указывать ссылки можно только на ресурсы внутри Вконтакте – например, на статьи, альбомы и пр. Ссылки на другие сайты добавить нельзя.
Теперь останется сохранить меню. В любое время вы можете отредактировать элементы или добавить новые.
ЗаключениеМы рассмотрели базовое оформление группы ВК – добавление аватара, обложки и меню. Как видите, в этом нет ничего сложного. Все действия выполняются в настройках сообщества на сайте или в приложении. Единственная трудность – создание самих изображений для элементов группы. Если у вас нет нужных навыков или не хотите тратить на это время, лучше доверить работу профессионалу.
Вам понравилась статья? 2 0
Руководство по оформлению группы в соцсети «ВКонтакте»
БАЗОВОЕ ОФОРМЛЕНИЕ: блоки и элементы
Создание аватарки
Работа над обложкой
Кнопки активного действия
Настройка меню
Создание картинки товаров
Закрепленный пост
Работа с виджетами
Блок с видеозаписями
Раздел со статьями
Блок со ссылками
Раздел с картами
Оформление подкастов
Блок с обсуждениями
Как оформлять посты?
ФОРМАТЫ ИЗОБРАЖЕНИЙ
Полезные сервисы для дизайна группы в «ВК»
ЗАКЛЮЧЕНИЕ
ВАЖНО! Если паблик оформлен непрофессионально, то клиенты вполне логично могут сделать вывод, что вы к своей работе относитесь так же. Поэтому важно позаботиться, чтобы страница «ВКонтакте» была красивой, аккуратной и удобной в использовании.
Грамотно продуманный дизайн группы VK покажет потенциальным клиентам, что вы серьезная компания, с которой можно и нужно работать. При этом правильно сделанная страница поможет пользователям сориентироваться и найти нужную информацию без лишних звонков и переписок.
Хотим сказать сразу, что универсальных рецептов по созданию красивой группы «ВКонтакте» нет. При этом сообщество ставит свои рамки: есть набор блоков и элементов, а также требования к размерам изображений. Поэтому процесс работы над оформлением паблика не такой легкий, как может показаться на первых порах. Рекомендуем вам прочитать эту статью до конца, чтобы узнать все возможности платформы и использовать их по максимуму.
БАЗОВОЕ ОФОРМЛЕНИЕ: блоки и элементы
При оформлении группы VK необходимо учитывать цели, задачи и особенности вашего бизнеса на площадке. Чтобы сообщество выглядело полноценным, кому-то достаточно использовать минимум блоков и элементов, другой же компании понадобится разработать полный пакет, включая шаблоны товаров и постов. В этой главе мы пройдемся по всем пунктам, чтобы вы понимали полный спектр возможностей «ВКонтакте».
Создание аватарки
Единственный элемент, который виден юзерам еще до момента входа в группу, – это аватарка. Она также является главной картинкой паблика. Логично, что от привлекательности этого элемента зависит, зайдет пользователь в сообщество или нет. Поэтому оформлению нужно уделить особое внимание.
Что ставить на аватарку? Есть несколько проверенных вариантов:
- анонс нового продукта, услуги или события;
- акции, распродажи или скидки;
- преимущества компании;
- тематические картинки;
- название компании;
- логотип.
Важно, чтобы текст можно было прочитать, поэтому делайте его достаточно крупным. Зрителям должно быть понятно, что на картинке. Кстати, не рекомендуем брать изображения со стоков, они не вызывают доверия пользователей.
Чтобы аватарка не затерялась среди прочих, сделайте ее яркой, запоминающейся и необычной. Подумайте, на что вы сами обращаете внимание? Способов выделиться очень много, просто нужно задаться этим вопросом и проявить немного креативности.
СОВЕТ. Хотите, чтобы аватарка выглядела современно? Тогда сделайте ее в стиле минимализм. Сейчас в тренде максимально простое и аккуратное оформление.
Подумайте, какой цели вы хотите добиться с помощью аватарки, и отталкивайтесь от этого при ее создании!
Работа над обложкой
Когда пользователь переходит в группу, первое, что бросается ему в глаза, – это обложка. Она представляет собой горизонтальный баннер в верхней части страницы. Главная ее задача − отражать тематику и род деятельности компании. Логичнее всего использовать обложку таким образом, чтобы привлечь внимание юзеров и дать им самую важную и цепляющую информацию.
ВАЖНО! В десктопной и мобильной версиях «ВКонтакте» размеры обложек отличаются. Со смартфонов люди увидят только центральную часть картинки. Выбирая дизайн, учитывайте этот момент.
Обложка может быть трех видов.
- Статичная. Представляет собой изображение без анимации. Создавая креатив, вы можете добавить в него все что угодно: начиная с названия и девиза вашей компании, заканчивая всевозможными акциями, предложениями и даже конкурсами. Немаловажно упомянуть контакты, УТП, адрес и призыв к действию.
- Динамичная. Это тоже картинка, но с интерактивными элементами, которые привлекают внимание, а иногда даже подкупают подписчиков. Такими элементами могут быть: топ активных пользователей, курсы валют, ники недавно вступивших пользователей, прогноз погоды или счетчики чего угодно. Динамический баннер на обложке повысит вовлеченность и стимулирует общение аудитории.
- Живая. Относительно недавно оформить группу в ВК стало возможно и с помощью живой обложки с видео или анимацией. В такой баннер-обложку можно загрузить максимум 5 картинок или небольших видео, которые будут сменяться автоматически. Но просматривать такую обложку получится только с мобильного телефона. В десктопном варианте баннер будет статичным, отображая одну заданную картинку. Такие обложки крупнее обычных в несколько раз и раскрываются почти на весь экран. Видео можно показывать со звуком.
Рекомендуем протестировать разные форматы обложек и посмотреть, какие из них получат больший отклик у подписчиков и новых пользователей.
Кнопки активного действия
Следующими элементами, визуально продолжающими шапку, являются кнопки действия. Они подводят пользователя к тому действию, которое вы ждете. Хотите продаж – направляйте сразу к товару или услуге, хотите больше подписчиков – приглашайте вступить в сообщество.
Примеры названий для кнопок активного действия:
- «Зарегистрироваться на вебинар»;
- «Подписаться на сообщество»;
- «Подписаться на рассылку»;
- «Узнать подробности»;
- «Перейти на сайт»;
- «Купить продукт»;
- «Задать вопрос»;
- «Позвонить».
Настраивать название и функционал кнопок можно в разделе «Управление».
Настройка меню
Меню является навигатором, облегчая поиск информации для подписчиков. Наличие меню необязательно, но оно поможет вам сделать группу более легкой для восприятия. Это особенно актуально, если она создана для продаж или демонстрации товаров и услуг.
Задача меню − перенаправлять пользователей в основные разделы группы. У каждой компании разделы будут разными, мы остановимся именно на расположении меню и на том, как оно может выглядеть.
«ВКонтакте» можно создать меню четырех видов.
- Виджет-меню. Представляет собой миниатюрные иконки под названием или описанием сообщества. Настройка осуществляется в разделе «Управление сообществом». Важно понимать, что при использовании этого варианта переход на внешние ссылки будет невозможен.
- Закрытое. В этом случае меню будет видно только при нажатии на вкладку, которая размещается в блоке «Свежие новости» на главной странице. В дальнейшем блок можно переименовать.
- Открытое. Это отдельная вики-страница с возможностью настройки переходов между разделами. Чтобы гости или фолловеры могли зайти в меню, необходимо будет создать закрепленную запись с кнопкой «Открыть меню».
- Скрытое. Разворачивается только при нажатии на активную ссылку в описании группы. Оформить ссылку вы можете в виде текста или эмоджи.
Виджет-меню − это наиболее популярный вариант. Немало групп используют открытый тип, считая, что чем меньше кликов до полезной информации, тем лучше. Некоторые сообщества и вовсе предпочитают обходиться без меню, но они, как правило, некоммерческие.
Создание картинки товаров
Немаловажная часть любой коммерческой группы − товары. Товарная галерея на платформе «ВКонтакте» безразмерная, вы можете размещать сотню и даже тысячу товаров. Картинки товаров в центральной части экрана размещают с помощью виджета, размещать их можно и в правом меню. При большом количестве товаров их чаще группируют в фотоальбомы. Это удобнее, так как юзер, кликнув на альбом, сразу увидит перед собой целую товарную линейку.
Закрепленный пост
Закрепленный пост демонстрируется над лентой публикаций. Часто именно в нем собрана самая важная информация о компании, продуктах, УТП и ключевых преимуществах. Его можно использовать, чтобы быстро донести до пользователей важную информацию. Например, расскажите в посте про анонс мероприятия, назовите дату вебинара или сроки поступления новинок в интернет-магазин. В этом случае ваша цель – проинформировать подписчиков и подогреть их интерес.
Если вы только создали группу, то логично будет закрепить пост-знакомство. Расскажите о компании, продемонстрируйте свое уникальное торговое предложение и ключевые преимущества. Это можно сделать не только в формате текста с изображениями, но и в виде презентационного видеоролика.
Закрепить можно и продающий пост. Это будет эффективно, если у вас, например, запланирован старт продаж онлайн-курса. Такая публикация должна содержать оффер для целевой аудитории и триггеры, которые ускорят процесс принятия решения о покупке.
Как еще можно использовать закрепленный пост? Некоторые компании размещают в нем ссылку на лид-форму. Такой вариант помогает собрать заявки и информацию о потенциальных клиентах. Нередко в закрепленных публикациях устраивают различные активности с возможностью получить подарки или бонусы.
Работа с виджетами
Виджеты «ВК» − это дополнительные блоки, в которых можно размещать важную информацию для подписчиков или функциональные элементы. Они помогают в коммуникации с аудиторией и облегчают работу администраторов. Важными виджетами являются:
- Приветствие. Помогает создать персонализированное обращение к каждому подписчику. Расположено над новостной лентой, видно как в десктопной, так и в мобильной версии «ВКонтакте». Обращение по имени − отличный повод привлечь внимание человека и сделать его более лояльным. В виджет-приветствии можно замотивировать пользователя сделать какое-либо целевое действие. Например, подписаться на рассылку, принять участие в конкурсе, почитать отзывы, перейти в другой раздел или написать в Директ.
- Оформление заявки. Подходит для повышения уровня продаж. Это встроенная лид-форма от сервиса «ВК», которая позволяет настроить поля так, как вам нужно, и получить заявки и информацию о клиентах, не заставляя их переходить на сайт или писать в Директ. Заявки с виджета могут копиться в специальной структурированной табличке или попадать в ваши личные сообщения.
- Сбор отзывов. Товары или услуги со множеством положительных отзывов вызывают больше доверия у пользователей. Поэтому, если вы собираете обратную связь от покупателей, покажите ее на всеобщее обозрение с помощью виджета. Такой прием повысит не только продажи, но и лояльность аудитории к продукту, а также количество фидбек-комментариев.
Виджеты − это инструменты, которые помогут донести нужную информацию до подписчиков. Но крайне важно определить, что именно вы хотите рассказать и показать потенциальным клиентам. Исходя из этого, и добавляйте те или иные виджеты.
Блок с видеозаписями
В группе «ВКонтакте» видео можно размещать в сайдбарах или в центральной части экрана. Пользователь в первую очередь видит именно обложки роликов. Поэтому важно сделать их красивыми и выдержать в едином стиле.
Раздел со статьями
Статьи, оформленные с помощью вики-разметки, можно размещать в правом меню или в центральной части страницы над лентой новостей. Второй вариант считается более заметным и, как правило, приносит большее количество кликов. Для каждой из статей важно подобрать красивые обложки, потому что, как и в случае с видеозаписями, подписчики будут видеть их в первую очередь. Рекомендуем создавать обложки в едином стиле, это улучшит визуальное восприятие блока.
Блок со ссылками
Для добавления ссылок необходимо создать специальный раздел, который будет располагаться в правом боковом меню. Считается, что этот блок помогает улучшить коммуникацию с пользователями, так как в нем они могут найти несколько вариантов для связи и общения с компанией.
Какие ссылки размещать? Это могут быть ссылки на другие соцсети, сайт, внешний блог, ВК-сообщества партнеров и отзывы. Если группа создана для развития личного бренда, то будет логичным указать ссылки на другие страницы этого человека.
Раздел с картами
Карта поможет визуализировать ваше местоположение. Это актуально, если речь о локальном бизнесе с геопривязкой или о бизнесе, у которого есть точки продаж или офис в офлайне.
Оформление подкастов
В последнее время все большую популярность набирают подкасты. Хорошо, что платформа поддерживает этот формат. Подкасты в VK представляют собой аудиоблоги с расширенными возможностями прослушивания: нет рекламы, есть фоновый режим, можно замедлять или ускорять аудиодорожку. При публикации можно добавлять текст, изображения, видео и опросы. Разместить подкаст можно над лентой или сбоку.
Блок с обсуждениями
Люди в соцсетях любят участвовать в обсуждениях, так что этот блок в любом случае будет полезен. Обсуждения располагаются в правом боковом меню. Если вам необходимо собирать мнения и впечатления о продукте, то сделайте раздел «Отзывы». Отвечать на вопросы пользователей будет удобно, если создать категорию «Вопрос – ответ». Рекомендуем опубликовать «Правила сообщества» и рассказать там, какие посты и комментарии разрешены к публикации, а какие будут удаляться модераторами. В дальнейшем при возникновении недопониманий с подписчиками вы сможете ссылаться на эти материалы.
Как оформлять посты?
Дизайн записей на стене должен быть в едином стиле. Это нужно для повышения узнаваемости вашего бренда и формирования доверия к нему. Но необязательно использовать один шаблон. Допустимо применять различные подложки для разных видов рубрик. Например, для информационных постов – одна подложка, для развлекательных – другая.
Самый распространенный вариант постов – это картинка или видео с текстом. Этот формат подойдет для любого контента. Для развлечения или информирования пользователей используйте аудио или плейлисты. Например, в группе по фитнесу можно выложить музыку для тренировок, а в паблике по организации мероприятий − подборку песен для какого-либо праздника. Периодически можно проводить анонимные и общедоступные опросы, они повышают активность.
ФОРМАТЫ ИЗОБРАЖЕНИЙ
Группа должна одинаково круто смотреться как на ПК, так и на смартфоне. Этого можно достичь, если адаптировать содержимое под просмотр на всех устройствах. Для этого при оформлении страницы необходимо подбирать форматы изображений. «ВКонтакте» предъявляет следующие требования:
- обложка для видеозаписей выставляется автоматически, но ее также можно загрузить, размер в этом случае должен быть 1280 х 720 пикселей;
- для карточек товаров подойдет любой квадратный формат;
- файлы, загружаемые в фотоальбом, – 1200 х 800;
- изображения для поста − 510 х 510;
- заставка для статьи − 510 х 286;
- обложка − 1590 х 400;
- аватар − 200 х 200.
Полезные сервисы для дизайна группы в «ВК»
Для оформления паблика «ВКонтакте» необязательно прибегать к помощи профессиональных дизайнеров и платить за это деньги. Конечно, ничто не заменит «Фотошоп», если нужно сделать полноценную ретушь или сложный коллаж, но для простых вариантов оформления может хватить и функционала онлайн-редакторов. Мы рекомендуем вам протестировать следующие программы:
- Canva − популярный редактор изображений, в котором много готовых шаблонов;
- Adobe Spark − приложение, в котором можно загружать шрифты и брать готовые шаблоны;
- Pixabay и Pexels − бесплатные фотостоки, которые подходят для поиска иллюстраций и фото;
- FontStorage – хранилище бесплатных шрифтов;
- SMMplanner − сервис отложенного постинга.
Используйте эти сервисы, чтобы оформить свою группу «ВКонтакте» быстро и красиво.
ЗАКЛЮЧЕНИЕ
Как мы говорили в начале статьи, продуманный дизайн группы «ВКонтакте» покажет потенциальным клиентам, что вы серьезная компания, с которой можно и нужно работать. Сейчас не нужно нанимать дизайнеров для разработки визуала, все материалы можно создать самостоятельно с помощью простых онлайн-конструкторов. Главное − начать. Удачи!
социальная сеть, мессенджер в App Store
Описание
ВКонтакте объединяет десятки миллионов людей, предлагая неограниченные возможности для общения, знакомств, развлечений, бизнеса и обмена новостями из любой точки мира. В приложении можно слушать музыку, смотреть видео и клипы, следить за своим здоровьем, играть в игры и делать покупки.
Оставайтесь на связи с друзьями и семьей. В мессенджере можно общаться в групповых чатах и личных сообщениях как с друзьями ВКонтакте, так и с людьми из контактов вашего телефона. Объединяйте неограниченное количество людей в видеозвонках бесплатно и без ограничений по времени.
ВКонтакте есть решения для самых разных повседневных ситуаций:
— Откройте для себя все, что может предложить одна из крупнейших социальных сетей в мире. Встречайте новых друзей и присоединяйтесь к онлайн-сообществам. Оставайтесь рядом с далекими друзьями с помощью мессенджера и звонков ВКонтакте.
— Слушайте любимую музыку и легко находите новые любимые песни благодаря персональным рекомендациям.
— Смотрите и создавайте клипы ВКонтакте, короткие вертикальные видеоролики на самые разные темы.
— Наслаждайтесь прямыми трансляциями, делитесь фотографиями, играйте в игры и читайте интересные новости в тематических лентах.
— Узнавайте что-то новое в подкастах и загружайте свои.
— Следите за своим здоровьем и оставайтесь в форме. Используйте ВКонтакте Здоровье, чтобы соревноваться с друзьями и посмотреть, кто может ходить больше всех. Ваши шаги учитываются благодаря Apple Health. Так что совершите долгую прогулку и попытайтесь добраться до вершины таблицы лидеров!
Условия использования: vk.com/terms
Политика конфиденциальности: vk.com/privacy
000Z» aria-label=»3 March 2023″> 3 марта 2023 г.
Версия 8.19.1
Мы добавили новые значки приложений, в том числе весенние капли дождя, нежные цветы, стиль комиксов и некоторые другие. Вы можете выбрать любой из них и включить его в настройках внешнего вида вашего приложения.
Рейтинги и обзоры
149 оценок
помощь
я уже зарегистрирован в вк, но когда я попытался войти, он не пускает меня и говорит, что кто-то уже вошел в систему с моей учетной записью
Сбой сервера
Я не могу войти в свою игру через ВКонтакте, а мне постоянно пишет, что «не могу войти под чужой учетной записью». Всегда так было после обновления.
Почта
Мне это очень не нравится, потому что я уже предлагал какой-то пост и не мог удалить свой пост из этой группы!
Разработчик, ООО «В Контакте», указал, что политика конфиденциальности приложения может включать обработку данных, как описано ниже. Для получения дополнительной информации см. политику конфиденциальности разработчика.
Данные, используемые для отслеживания вас
Следующие данные могут использоваться для отслеживания вас в приложениях и на веб-сайтах, принадлежащих другим компаниям:
- Контактная информация
- Идентификаторы
Данные, связанные с вами
Следующие данные могут быть собраны и связаны с вашей личностью:
- Покупки
- Расположение
- Контактная информация
- Контакты
- Пользовательский контент
- История поиска
- История браузера
- Идентификаторы
Данные, не связанные с вами
Могут быть собраны следующие данные, но они не связаны с вашей личностью:
- Здоровье и фитнес
- Пользовательский контент
- Идентификаторы
- Данные об использовании
- Диагностика
Методы обеспечения конфиденциальности могут различаться в зависимости, например, от используемых вами функций или вашего возраста. Узнать больше
Информация
- Поставщик
- В Контакте ООО
- Размер
- 301,6 МБ
- Категория
- Социальная сеть
- Возрастной рейтинг
- 12+ Нечастые/мягкие зрелые/наводящие на размышления темы Нечастая/умеренная ненормативная лексика или грубый юмор Редкий/умеренный сексуальный контент и нагота Нечастое/мягкое реалистичное насилие Нечастое/умеренное насилие в мультфильмах или фэнтези Нечастые/мягкие темы ужасов/страха
- Авторское право
- © 2023 ВК
- Цена
- Бесплатно
- Сайт разработчика
- Тех. поддержка
- политика конфиденциальности
Опоры
Еще от этого разработчика
Вам также может понравиться
стикеров ВК на продажу | Redbubble
Tags:
tank, german, combat vehicle, vk 4501, army, heavy tank, vk4502, military, panzerkampfwagen vi, pzkpfw vi
Tiger I Sticker
By deathdagger
From $1.14
Tags:
Holden, vl, commodore, turbo, calais, red, rb30, car, auto, race, walkinshaw, дозы, 1987, 1988, вн, вк, австралия, руководство, автомобильный, транспортное средство, bt1, полицейский, перехватчик, чейзер
Holden VL Commodore Turbo Badge Sticker
larue, victoria, schwab, ve, ve, arc, vk, special edition, devil, goodreads, fan art, book, invinsible life, adeline, pack, забытый, одинокий
цитаты addie larue The Invisible Life of Addie LaRue Наклейка
Автор krypton4shirt
От $1. 38
Метки:
холден, ford, hsv, vk commodore, vk group a, vk ss, холден вк, батерст, commodore, vl turbo, Holden v8, aussie v8, v8, monaro, maloo, clubsport, barra
Холден ВК СС Группа А (синяя) Наклейка
By mtsala illustrations
От $1.33
Теги:
София Карсон потомки, потомки София Карсон, мал, потомки, потомки 3, мал потомки, потомки, мал, голубь камерон софия карсон, любовь, фиолетовый, синий, панк, вау, девочка, девочки, малыш, дети, для детей, для девочки, принцесса, милая, милая, королева мал, потомки голубь камерон, потомки голубя камерона, голубь камерон мал, вк
Evie Queen — Потомки 3 Наклейка
By Magical Forest
От $1.18
Теги:
Бен и Мал, потомки Софии Карсон, мал и Бен, мал, потомки, потомки 3, мал потомки, потомки мал, голубь Кэмерон , софия карсон, любовь, фиолетовый, синий, панк, вау, девушка, девочки, малыш, дети, для детей, для девочки, принцесса, милая, милая, королева мал, потомки голубь камерон, потомки голубя камерона, голубь камерон мал, вк , cuple, романтический cuple, романтика, летняя любовь, лучший cuple, потомки 2, потомки mal и ben, королева и король, принцесса и принц, закат, солнечный свет, солнце
Mal and Ben Romantic Summer Love — Descendants 2 Наклейка
By Magical Forest
От $1. 18
Метки:
club, vk, vl, Holden, commodore, car
Club VK-VL Calais By 90j005 And 90j005 90j005 Наклейка 90ma005 And 90ma005
From $1.28
Теги:
танк, немецкий, боевая машина, vk 4501, армия, тяжелый танк, vk4502, военный, panzerkampfwagen vi, pzkpfw vi0005
Теги:
аид, потомки, д3, дисней, мал, малефисента, вк, гранж, синий, угли
Делай то, что должен делать — Наклейка Аида D3
питер, брок, холден, коммодор, вк, команда, hdt, гонки, v8, суперкары, автоспорт, туринг, автомобили, atcc, marlboro, батерст $1.58
Теги:
австралийские автомобили, холден, вк, вк коммодор, коммодор, австралийский
VK Commodore Sticker
от Ozhumour
от $ 1,28
Теги:
Sesh, UNI, VK, Vodka, WKD, Университет, Bournemouth, Clubbing, Allak
VK -VK -Vodka Fulaved By Shesh Strawer Strawer Strawer Strawer Strafor Strawer St. Strafor St. Strafor St. Strafor St. Strafor St. Strafor St. Strafor St. Strafor St. Strafor St. Strafr.
из $ 1,18
Теги:
Jrock, Visual Kei, J, Mana, Juka, Seth, K, Malice Mizer, VK, Vkei, Mana Sama
Moi Dix Mois
By Insidebbeast
от $ 1,18
9
9
9
9
996699
9
996699
96669
69
69
69
6
69
9
. Теги:
visual kei, vk, v系, gackt
gackt Наклейка
By rosarie-s
От $1.18
Метки:
gascoigne, holden, commodore, vk, peter, brockaustralia , авто, гонка, белый, красный, скорость, быстро, винтаж, исторический, мт, панорама, батерст, 1000, шина, колесо, драйв, быстро, скорость, быстро, победа, победитель, гонка, бензин, резина, мальборо, 22 , ретро, vl
ВКонтакте Brock Edition Commodore Sticker
By Andre Gascoigne
From $1. 48
Метки:
сс, Holden, commodore, vc, vk, vl, vn, vs, vt, vx, vy, vz, ve, vf, car, australia
SS Наклейка владельца
By Andmaj
Из $ 1,28
Теги:
Потомки, В.К., Карлосдевилл, Гранж, Брызги, Дисней, Роттен до ядра
Десконец В.К.
vk
Семейная наклейка Psomas
от Victoriakahili
из $ 1,18
Теги:
Blade Runner, Voueth Kampff, Harrison Ford, Nexus6, Nexus 6, Vk Test, Replicant, Rachael
Voight -Campf
Теги:
цветной карандаш, фотореалистичный, gmh, Holden, commodore, groupa, blue meanie, Muscle Car, автомобильный, классический автомобиль, автомобиль, автомобили
1984 VK Holden Commodore SS Group A Sticker
By Wayne Sotogi
От $2,07
Метки:
visual kei, thegazette, vkei, vk, japan, japanese, japanese music, jrock, jpop, black butler, uruha, ruki, aoi, reita, kai, yune , группа, японский металл, jmetal, металл, логотип
The GazettE Logo Sticker
брок, commodore ss, ss commodore group a, ss commodore, brock group a, blue brock commodore, hdt group a, hdt vk group a, hdt blue meanie, 1985 brock group a, 1985 vk group a
VK Group A «Blue Meanie» Наклейка
By CuccoviaDesign
От $1. 38
military, panzerkampfwagen vi, pzkpfw vi
Tiger I Sticker
By deathdagger
From $1.14
Tags:
vk, ss, commodore, holden, v8, peter brock, burnout
VK SS Commodore decal Sticker
By майкон
Из $ 1,28
Теги:
VK, напиток, водка, студент, Sesh, Night Club, Cameo, Wkd
VK Drink Vodka Student Sesh Sticker
By Markey1998
от $ 1,18
TAGS1998
от $ 1,18
666666666666666666666666666666666666666666666666666666666666666666666. , commodore, автомобиль, vk vl, vn, vs, vt, vx, vy, vz, ve, vf
Кале Life Sticker
By Andmaj
От $1.28
Теги:
ума, потомки, потомки ума, ума потомки 3, потомки 3 ума, потомки 3, china anne mcclaine, china mcclain, ума чина, ума чина энн макклейн, софия карсон, голубь камерон, черный, черная девушка, черные дети, черные девушки, девушка, девушка, черная принцесса, принцесса , королева, девочка принцесса, для девушки, женщина, женщины, любовь, милая, милая, красивая, удивительная, голубая, блеск, ночь, вк
Принцесса Ума — Потомки 3 Наклейка
By Magical Forest
От $1. 18
Метки:
long, live, evil, гнилой, ядро, потомки, 2, 3, мал, малефисент, силуэт, спящая, красавица, эви , carlos, jay, uma, disney, disney world, disneyland, disneybound, disneybound, волшебное королевство, злодеи, злодей, vk
Evil Icon Sticker
, v系, 悪の華, бак тик, черный, cd
aku no hana CD Наклейка
By rosarie-s
From $1.18
Tags:
brock, vk, ss, blue, car, Holden, hsv, commodore, vl
And BROCK9 VK0 StickerИз $ 1,28
Теги:
Descendants3, Jay, Jafar, VK, Descendants
Good to Bad Eeshetic — Jay Sticker
от Jaila Desper
от $ 1,18
. , visual kei, vk, jrock, chibi, cute, kawaii
Quarentine Uruha Sticker
By galleonsgoodies
От $1. 18
Метки:
commodore, Holden, vl, car, turbo, australia, v8, gmh, ss, hsv, ve, vns, vf, vz, автомобильная , vs, Walkinshaw, Bathurst, мощность, vl turbo, bt1, vk, белый, gts, aussie, ssv, vt, clubsport, vy
VL Walkinshaw SS Group A Sticker
By Chrome and Coils
От $1,88
Теги :
коммодор, вагоны, коммодор вагоны, холден, холден коммодор, холден вагон, автомобиль, автомобили, сс, сс коммодор, австралия, быстрые автомобили, трамваи, длинная крыша, длинная крыша, ве, вф, вз, ви, вк, вн, вл
Commodore Wagons V1 (Белый) Наклейка
Автор ArtStinkInk VK, VL, VN, CAR, Australia
Blue VZ Сенатор сенатор
от Andmj
от $ 1,28
Tags:
VK
Старший Стейнер
9016. , thegazette,gazeto, thegazette, visual kei, vk, visualkei, jrock, kawaii, chibi, cute, lamen, ramenGazefoodies: стикер Reita’s Ramen
By galleonsgoodies
От $1.18
Теги:
slr, 5000, torana, Holden, Ford, Muscle, a9x, gthoout, falcon, vk, commod, vf, vf
Наклейка Torana SLR 5000
От ClassicAuto
От $1.28
Теги:
vk, vaca key, florida, fl, vaca key florida, florida keys, овал, овал, вк овал, евростиль, евробампер стиль, dodgem design, dodgemdesigns
VK — Vaca Key Florida Oval Design Sticker
от Dodgemdesigns
от $ 1,23
Tags:
Holden, VK, Commodore, BT1, Yellow, Car, Holden Commodore, GMH
Holden VK St. St. St. St. St. St. St. St. St. St. St. St. Stractemaster
6666966696669606
9
9
9
9
9
9
9
9
9
9
9
9
9
9
9
9.
From $1.28
Метки:
jrock, visual kei, французская аристократия, французские аристократы аристократы, версаль, вк, японец, камиджо, хизаки, теру, масаси, юки
Versailles Sticker
5
5
From $1.18
Теги:
Gazeto, thegazette, thegazette, uruha, jrock, visual kei, vk, visualkei, bubbletea, пузырьковый чай, милый, каваи, чиби
От $1.18
Теги:
the invisible, addie, larue, victoria, schwab, ve, ve, arc, vk, special edition, devil, goodreads, fan art, book, invinsible life, adeline, pack
addie Ларю цитирует Невидимая жизнь Адди ЛаРю Наклейка
By krypton4shirt
From $1. 38
Tags:
vk, vl, brock, commodore, holden
Australia v8 muscle car VK VL Sticker
By concuido
From $1.53
Tags:
vk
Angel’s Наклейка на день рождения
By VictoriaKahili
От 1,18 $
Теги:
Коллекции vk
Коллекции Vk Art Season
By Vishal Kumar
От 1,18 $
Теги:
Trio Thetrio Hannah Victoria Julia Sketch VK
The Trio Sticker
от Victoriakahili
из $ 1,18
Tags:
666, addiab, viceab, viceab, viceab, viceab, viceab, viceab, viceab, viceab, viceab, vic , специальное издание, дьявол, goodreads, fan art, book, invinsible life, adeline, pack, забытый, одинокий, пять, рейтинг, мудрый, хороший
Цитаты адди Ларю Невидимая жизнь Эдди ЛаРу Наклейка
$1,38
Теги:
социальные, СМИ, серьезно, ущерб, ваше, психическое, здоровье, поврежденный, ущерб, серьезный, серьезный, серьезно, facebook, instagram, Snapchat, vk, whatsapp, телефон, электронная почта, текст, черный, белый, правда, цитата, реальная, депрессия, тревога, грусть, одиночество, депрессия, грусть, плохо
Социальные сети серьезно вредят вашему психическому здоровью vk 4501, армия, тяжелый танк, vk4502, военные, panzerkampfwagen vi, pzkpfw vi
Stuh 42 Sticker
от DeathDagger
из $ 1,14
Tags:
VK, Roma, UM
мм . .. Рома (VK) Наклейка
966 от $ 1,28
6669
9
. , kawaii, thegazette, visualkei, vk, thegazette, ruki, jrock, japan, japanese, minimalist
Наклейка Smol Fashion Bun Sticker
By Gabi Sardenberg
От $1,18
Теги:
научная фантастика , фильм, кино, фильмы, компьютеры, технология, программирование, программист, игра, гейминг, хак, взлом, хакер
VK Sticker
By jpjmoffett
From $1.18
Tags:
die goldene emma, gala, goldene emma, goldene emma 2022, pietsmiet emma, pietsmiet gala, pietsmiet goldene emma, pietsmiet preis, pietsmiet preisverleihung, pietsmiet react, preisverleihung, реагировать, josylvio, ездить или умереть, эско, beats by esko, ma3seb, 2 gezichten, рэп, трэп, хип-хоп, клуб, регги, дэнсхолл, ван класс, вк, вк весь день, вк сезон, вк город, хелла cash gang, almere, amsterdam, neo magazin royale, neo magazin, zdf, zdf magazin
SKATE OR DIE Наклейка
Магазин Rastko
От $1,18
Метки:
calais, holden, commodore, car, vk, vl, vn, vs, vt, vx, vy, vz, ve, vf
Calais List Sticker
By Andmaj
От $1,28
Теги:
dir en grey, visual kei, vk, jrock, japanese, j, kyo, kaoru, Shinya, die, toshiya, japanese, music, bands, heavy metal
Серо-красный | Наклейка со списком участников группы
от LolirotSav
From $1. 23
Метки:
red, calais, car, Holden, commodore, australia, andmaj, vx, vt, vy, vz, ve, vf, vs, vl, vk, vn
Красная наклейка VX Calais
от Andmaj
Из $ 1,28
Теги:
Kuroyume, 黒 夢, VK, Japan
黒 夢 Kuroyume Sticker
,
от $ 1,18
. , зин, visual kei, jrock, vk, kuze, daisuke, rucy, mashashi, yuki, japanese, versailles philharmonic quintet
Jupiter 2020 Sticker
By InsideBeast
From $1.18
Tags:
phil lester vk telegram
Phil Lester VK Telegram Sticker Sticker
By teepyshop
From $1.18
Tags:
descendants3, carlos de vil , vk, Crusela de vil, grunge, spatter, disney
Good to be Bad — Carlos De Vil Наклейка
By Jaila Desper
От $1. 18
, thegazette kai, jrock, vk, visual kei, visualkei, kawaii, пузырчатый чай, баблти
Gazefoodies: Чай с пузырьками Кая Наклейка
By galleonsgoodies
От $1.18
Метки:
Holden, hsv, vf, gts, gtsr, clubsport, commodore, ve, vz, vtv, vy, vx, vx vn, car, andmaj, australia
Son Of A Gun GTSR Sticker
By Andmaj
От $1.28
Метки:
танк, немецкий, боевая машина, vk 4501, армия, тяжелый танк, vk4502, военный, panzerkampfwa , pzkpfw vi
Тигр I Наклейка
от Deathdagger
от $ 1,14
VK Sticker
от OtterCreekcc
от $ 1,18
TAGS:
Taekook, Vkook, Tae, JK, V, VK
TAEKOK, Vkook, TAE, JK, V, VK
666666666666666666666666. От $ 1,58
Теги:
Visual Kei, J, VK, V Kei
Galace Visual Kei — Eternal Matina Sticker
By4r3a1r3m1x
из $ 1,18
Теги:
66666666666666.0005
Eat Sleep Bohurt VK Armored Full Contact Sticker
от Renatosanches
от $ 1,38
Теги:
Mejibray, SM, SM2, Slivers, Sliver Exe, Sliver, Mejibray, Mejibray, Vkei, Visui, VK
5505055055055055055055055050505505055050505050505505050505505050505cн5505050505050505050505050505. mejibray SM2 stickers НаклейкаBy goopybird
От $1.18
Метки:
thegazette, thegazette, Gazetto, jrock, vk, visual kei, chibi, kawaii, cute, japanese, japan Наклейка
9005 Smol Gzt СарденбергFrom $1.18
Теги:
Кай, The Gazette, Gazetto, visualkei, visual kei, vk, japan, japanese, cute, jrock, j, thegazettekai, thegazette kai, thegazette, chibi, kawaii
Kai’s No Look Byebye Sticker
от GalleonsGoodies
от $ 1,18
Теги:
Gackt, Twitter, твит, ужасный, проклятый, певец, Visual Kei, Vk, Malice Mizer, Japan
.
Теги:
коммодор, вагоны, коммодор вагоны, холден, холден коммодор, холден вагон, автомобиль, автомобили, сс, сс коммодор, австралия, быстрые автомобили, трамваи, длинная крыша, длинная крыша, ve, vf, vz, vy , vk, vn, vl
Наклейка Commodore Wagons V2 (белая)
Автор ArtStinkInk
От $1.28
hq, hj, hx, hz, vb, vc, vh, vk, commodore, kingswood, aussie, австралийский, мускулистый, автомобиль, быстрый, 6-цилиндровый, рядный 6, рядный 6, mac
Holden 202 Motor Sticker , маслкар, автомобиль, моторизация, световая живопись, светлая, симмонс, взорванный, наддув, седан
White Holden VK Commodore Sticker
By HoskingInd
От $1.28
Теги:
vanesakovse 0 0 0 0 9 , vanesa, 9
VK Sticker
от Vanesa Kovse
из $ 1,48
Теги:
Дизайн искусства цифрового искусства Photoshop Car Тату , fanart, fan art
Наклейка Evie/Sofia
Автор jennyslass
От $1. 18
Теги:
танк, немецкий, боевая машина, vk 4501, армия, тяжелый танк, vk4502, военный, vipzkampfkwagen, panzerkampfkwagen0005
Tiger I Sticker
от DeathDagger
от $ 1,18
Tags:
Танк, немецкий, боевой транспортный Deathdagger
из $ 1,14
Теги:
VK, Vibe Killers, проверка атмосфера, проверка атмосфера
Vibe Check Sticker
By Elvythevelvy
от 1,18
Теги:
66. , клыки, глаза, ужас, надпись
Умрите, собаки! Sticker
By AerachnaNagren
From $1.18
Tags:
holden, vk, commodore, ss, gm, car, australia, sedan, red
Crusing VK Sticker
By Andmaj
From $1. 28
Tags:
фрида кало, фрида, кало, мексиканка, женщина права, фрида кало абстракция, абстракция, лицо, феминистка, мексика, vk91, vk91art, vidka91, vk cases, vkcases, vk dezine, телефон, найди свою вещь, по запросу, батик, кейс, футляры, скины, обложка
Frida Kahlo Abstract Sticker
By Vk91art
От $1.23
Теги:
танк, немецкий, боевая машина, vk 4501, армия, тяжелый танк, vk4502, военный, panzerkampfwagen
от Deathdagger
от $ 1,14
Теги:
Visual Kei, VK, Teen, Tik Tok, Mrendy, Cool, Pinterest, ビジュアル 系, カッコイイ
Visual Kei Sticker
By Andiiandii
от $ 1,48
.
1984 VK Holden Commodore SS ‘Peter Brock’ Edition — Automotive Classics Sticker
By Heartwall
От $1. 28
Теги:
черный, vx, кале, холден, коммодор, Walkinshaw, австралия, автомобиль, vk, vl, vn, vs, vt, vy, vz, ve, vf
Black VX Calais On Walkys! Наклейка
Автор Andmaj
From $1.28
Метки:
commodore, универсалы, commodore wagons, холден, холден коммодор, холден универсал, автомобиль, автомобили, сс, сс коммодор, австралия, быстрые автомобили, трамваи, длинная крыша, longroof, ve, vf, vz, vy, vk, vn, vl
Commodore Wagons V2 (черный) Наклейка
Автор ArtStinkInk
От $1.28
mal
Хорошо быть плохим — Эви Стикер
By Jaila Desper
From $1.18
Tags:
lamborghini, car, beutiful, popular, yellow, fast, trending, vk, artist, stylish
Lamborghini car Sticker
By VIKRAM JANGID
From $1. 18
Теги:
потомки, королева, трое
Потомки Рожденные в ВК 3 Наклейка
By reevesprattx
От $1.33
Теги:
офелия, шекспир, девушка, гамлет, женщина, романтика, средний возраст, леди, озеро, фэнтези, зеленый, вероника касас, вк арт
Ofhelia Sticker
от Verónica Casas
от $ 1,18
Теги:
Монополия, Телеграмма, VK, поведение, гендлмен, персонаж, мужчина, Thumb
Monoply
696696696696669669669669669696696696696696696666696696696696666966696666666666666666666669669666666666666666666666666666666666666 Метки:
веселый, япония, visual kei, вк, японец, гара, юу, кеничи, тэцу, группы, nero
МЕРРИ | Наклейка со списком участников группы
0005
хобби, пз ви тигр п вк, 45 01 п
Хобби- танки! Наклейка Pz-VI Tiger P (VK 45. 01 P)
AliciaOConno
От $1.18
Метки:
dadaroma, i take, vk, visual kei, visualkei, jrock, japan
I take Наклейка
5 kuroro
От $1.18
Метки:
танк тигр, вк 4501, тигр п, вк 45 01, пз ви, панцер, панцеркампфваген, танк второй мировой, тяжелый, военный, армейский танк, вар тандер, wot blitz, wotblitz, world of tank , Worldoftanks, Германия, Немецкий, Железный крест, вторая мировая война, Faawray, Игра, Любитель танков, Танковер, Танкер, Смешные военные, Масштабное моделирование, Модельер
Хобби- танки! Наклейка Pz-VI Tiger P (VK 45.01 P)
By FAawRay
От $1.67
Теги:
золотой жеребец, золотой, жеребец, феррари, лошадь, скачущий конь, арабская лошадь, скачки, красочный, типография, животное , Абстрактные, формула 1, Формула один, Enzo, Италия, Америка, Немецкий, Франция, Произведения искусства, Vk91, vk91art, Вк случаи, vk dezine, la fara, Скрипт vk, Мода, творчество, автогонки, абстрактное искусство, Moto gp , мексика, найди свою вещь, цвет, цвет, уникальный, классный
Золотой жеребец 1 Наклейка
By Vk91
От $1. 19
Теги:
киноварь, фэнтези, дракон, сокровища, золото, деньги, монеты, вероника касас, искусство vk Casas
From $1.18
Теги:
center side black Nine, centerside, blacknine, vk, lada, легенда для пацанов
CENTER SIDE BLACK NINE Наклейка
By Racingdecals
От $1.200005
Теги:
Bathurst, Holden, GM, VK, Commodore, Holden Commodore, Brock, Perkins, HDT
1984 Bathurst — Commodore VK Sticker
от Formulat
от $ 1,38
. я беру, visual kei, вк, тыква, чиби, япония
Halloween Volume 2018 Sticker
Курозеро
От $1.18
Теги:
орнаментальный, птица, сюрреалистический, сюрреализм, золото, черный, белый, природа, животное , вероника касас, вк арт, шикарная
Наклейка с декоративной птицей
Вероника Касас
От $1. 18
Теги:
visual kei, вк, дадарома, я беру, ангел, ангелы, бас, низкий, инструменты, человек, японский
0 Наклейка Dark Angel 5
0 Dark Angel By kurozero
От $1.18
Теги:
цветы, вк, карро, машина, синяя, эстетика, хиппи тачка, эстетичная машина
Вк с цветами Наклейка
By GeminisGem
От $1.18
Теги:0005
mejibray, visual kei, v kei, vk, группы, японский язык, япония, музыка
Mejibray | Наклейка со списком участников группы
Автор LolirotSav
От $1,23
Метки:
Holden, commodore, ve, ute, v8, supercars, supercar, boy, girl, ss, General Motors, maloo, morpheus, фиолетовый, autoart78, vb , vc, vh, vk, vn, vp, vx, vy, vr
Holden VE SS Ute Morpheus Purple Sticker
By Brad Van Geest
От $1. 58
Структура комплекса TnsB-транспозаза-ДНК типа V-K CRISPR- ассоциированный транспозон
ВВЕДЕНИЕ
Обнаружение адаптивной прокариотической иммунной системы под названием C Lustered R Egularly I Alindrom S HORT P Alindrom DASTERS ASERMIC DASTEATE (Alindrom HORT P Alindrom D. CRISPR-ассоциированные белки произвели революцию в науках о жизни. Системы CRISPR-Cas представляют собой весьма разнообразные комплексы рибонуклеопротеинов (РНП) с разным эволюционным происхождением. Они делятся на два класса, класс 1 и класс 2, первый из которых включает многосубъединичный эффекторный комплекс, а второй — единственный белковый эффектор 9.1259 1 . Два класса далее делятся на шесть типов (I-VI) в зависимости от идентичности нуклеазного модуля и множество подтипов в зависимости от того, какие другие белки Cas присутствуют в других функциональных модулях. В частности, члены класса 2 привлекли большое внимание, поскольку они были разработаны в универсальные РНК-управляемые нуклеазы для РНК-управляемого редактирования генома, что радикально изменило науки о жизни, позволив манипулировать геномом в живых организмах 2 .
Недавно было обнаружено, что несколько механизмов CRISPR-Cas связаны с Tn7-подобными системами транспозонов типов I, IV и V. Эти системы CAST 3,4 являются продуктом эволюционного процесса, в ходе которого транспозоны, подобные Tn 7 , рекрутировали систему CRISPR-Cas для мобилизации транспозонов. Эти комплексы не разрушают свою ДНК-мишень и действуют исключительно на прокариотах. Они вставляют большие ДНК-грузы (10–30 т.п.н.) в определенные области генома без необходимости гомологически направленной репарации 4,5,6,7 , сочетая точность выбора сайта CRISPR-Cas с интеграционными свойствами транспозонов 8 . Поэтому считается, что CAST представляют собой очень многообещающую систему для разработки инструментов редактирования генов следующего поколения.
CAST I-F, I-B и V-K подтипы, из Vibrio cholerae (vc), Anabaena variabilis (av) и Scytonema hofmannii (sh) соответственно, были обнаружены первыми 3,4 , но недавние биоинформационные поиски метагеномных баз данных значительно расширили известный репертуар CAST до более чем 1000 не повторяющихся подсистем, представляющих Типы I, IV и V 6 . На сегодняшний день все известные CAST происходят от Tn 7 -подобных транспозонов и включают соответствующие crРНК и генов Cas , необходимых для отбора мишеней 6,7 , и основной механизм транспозиции в Tn 7 -подобном локусе транспозона. Сюда входят гены TnsB, TnsC, TniQ (гомолог E. coli TnsD) и, в некоторых случаях, гены TnsA. По аналогии с системами транспозонов Tn7, белки CAST Tn 7 , как полагают, собираются в нуклеопротеиновый комплекс перед интеграцией, который включает TnsA (в типах I и IV), TnsB, TnsC и TniQ, чтобы регулировать транспозицию в инсерцию. сайт. TnsA — эндонуклеаза, расщепляющая 5′-концы транспозона 9.1259 9 и взаимодействует с TnsB, TnsC и ДНК 9,10,11 . TnsB является рекомбиназой и катализирует расщепление 3′-концов транспозона. В канонической системе Tn7 взаимодействие TnsA и TnsB необходимо для активации катализа 12 . TnsC является частью семейства AAA+ ATPase и направляет TnsA/TnsB к сайту встраивания 11,13 . Считается, что взаимодействие TniQ с ДНК-мишенью, связанной комплексом CRISPR-Cas, создает искажение в структуре ДНК, позволяя TnsC распознавать как TniQ, так и ДНК 9.1259 14 , что приводит к вставке транспозона в сайт прикрепления. Однако системы CAST типа V-K отличаются от Tn7 из-за отсутствия TnsA в их локусах. Поскольку CAST типа V-K не содержит какого-либо другого белка для замены эндонуклеазной активности TnsA, его транспозиция продуцирует коинтеграты, которые необходимо расщепить 15,16 . Разрешение коинтегратов, по-видимому, осуществляется посредством RecA-зависимой рекомбинации, как и в случае других транспозонов, лишенных фермента с 5′-эндонуклеазной активностью. Тем не менее, транспозоны из семейства Tn5053, которые также лишены TnsA, но содержат гомологи для TnsB, TnsC и TniQ, могут разрешать коинтеграты с использованием кодируемой транспозоном резольвазы TniR. Может ли резольваза, кодируемая хозяином, обеспечивать эту активность в типе V-K CAST, еще предстоит определить.
Транспозазы TnsB принадлежат к суперсемейству ретровирусных интеграз с каталитическим доменом рибонуклеазы H и мотивом активного сайта DDE. TnsB связывается с левым и правым концами транспозона и катализирует их расщепление с образованием свободных 3′-гидроксильных групп, которые позже используются в нуклеофильной атаке ниже целевой последовательности 17 . Наконец, он выполняет реакцию переноса цепи, которая приводит к вставке груза ДНК в целевой сайт.
Транспозаза shTnsB имеет гомологию с другими членами семейства интеграз DDE, такими как E. coli TnsB, MuA и Tn5053 (дополнительный рисунок 1). Структура E. coli (ec) TnsB в комплексе с концом транспозона предоставила новые доказательства, объясняющие различия в распознавании левого и правого концов элемента 18 . Однако идентичность последовательности ecTnsB с shTnsB низкая, что затрудняет понимание деталей механизма интеграции CAST и других систем транспозонов, содержащих интегразы семейства DDE.
Чтобы понять, как shTnsB способствует РНК-управляемой транспозиции в системе CAST V-K, мы определяем структуру комплекса shTnsB-ДНК, захваченного после реакции переноса цепи, с помощью крио-ЭМ одиночных частиц с разрешением 2,46 Å (рис. 1). Структура показывает запутанную архитектуру белка shTnsB вокруг ДНК, образующую псевдосимметричную сборку, в которой четыре субъединицы shTnsB могут быть сгруппированы в двух разных конформациях. ДНК в каталитических карманах DDE резко изгибается после реакции переноса цепи. Сайт-направленный мутагенез и анализы транспозиции in vivo выявили важную роль в транспозиции ключевых ДНК-связывающих остатков. Комплекс реакции переноса нити shTnsB открывает новые пути к пониманию управляемой РНК транспозиции в системах CAST.
Рис. 1: Крио-ЭМ структура субъединицы CAST shTnsB, представляющая посткаталитическое состояние комплекса переноса цепи. a Общая схема реакции интеграции транспозаз семейства TnsB. Активный центр DDE катализирует атаку H 2 O на концах транспозона. Прямая атака 3’ОН груза ДНК на сайт ДНК-мишени приводит к образованию комплекса переноса цепи (STC). Промежуточный продукт Шапиро разрешается репликацией, производящей дупликацию сайта-мишени и коинтеграцию 25 , которые могут быть разрешены с помощью RecA или резолвазы. b Архитектура белка shTnsB. c Карта плотности крио-ЭМ с глобальным разрешением 2,46 Å TnsB (STC) в посткаталитическом состоянии (см. Также дополнительные рисунки 6–7, дополнительную таблицу 2). Карта окрашена в соответствии с карикатурой, описывающей строение на панели d . д Мультфильм ШТнсБ-СТЦ. Комплекс состоит из 4 протомеров shTnsB и 6 олигонуклеотидов ДНК, представляющих посткаталитическое состояние STC (панель 9).1329 и ). С этой точки зрения целевая ДНК расположена вверху структуры, а более длинные концы транспозона расположены внизу. На схеме отмечены непереносимая цепь (NTS) и переносимая цепь (TS) транспозона. Пунктирные линии изображают взаимодействие между MD и NTD1 из-за кривизны ветвей ДНК. e Ленточная диаграмма, показывающая обзор сборки ШТнсБ-СТЦ.
Полноразмерное изображение
Результаты
Выделение shTnsB и создание комплекса переноса цепи (STC)
Рекомбинантный белок shTnsB был экспрессирован в E. coli и очищен с использованием комбинации аффинной и эксклюзионной хроматографии (SEC). Белок вел себя как мономер в SEC-MALS с молекулярной массой 68 кДа (дополнительные рис. 2a, b, методы). Мы проанализировали его ДНК-связывающие свойства с помощью EMSA с использованием олигонуклеотидов с разным количеством коротких и длинных концевых повторов (SR и LTR соответственно), присутствующих на левом и правом концах (LE и RE) последовательностей транспозонов (дополнительная рис. 2c, г, дополнительная таблица 1). Анализ сдвигов полос со всеми различными субстратами выявил лестницу дискретных полос, относительная подвижность которых обратно зависела от концентрации белка, предполагая связывание одного белкового мономера на повтор. Сборки выше шести или семи белков не наблюдались, поскольку их размер препятствовал попаданию в акриламидный гель (дополнительный рисунок 2c). Затем мы проверили, может ли shTnsB независимо связывать RE или LE. Оба комплекса, shTnsB-RE и shTnsB-LE, были обнаружены, что указывает на то, что присутствие обоих концов не требуется для связывания shTnsB с повторами (дополнительный рисунок 2d). Точно так же количество полос, наблюдаемых при смешивании shTnsB с RE, соответствует 5 сайтам связывания TnsB. Однако количество полос, обнаруженных при смешивании shTnsB с LE, составило 4, т.е. на одну больше, чем ожидаемые три сайта связывания. Это можно объяснить взаимодействием двух комплексов shTnsB:LE, подобным тому, которое наблюдается в комплексе STC (рис. 1), или в других структурах транспозазы, не связанных с ДНК-мишенью, например, в Tn5 19 . Однако агрегация комплекса shTnsB:LE также может вызывать сдвиг полосы и не может быть исключена как возможность. Наконец, мы проверили связывание shTnsB с двухцепочечной ДНК, содержащей только LTR (6)-SR (1) (дополнительный рисунок 2e), и, как и ожидалось, обнаружили две полосы, соответствующие двум сайтам связывания shTnsB. Кроме того, мы инкубировали этот комплекс в буфере, содержащем различные концентрации NaCl, при 37 и 45 °C, чтобы определить, могут ли эти переменные изменить аффинность shTnsB. Однако на ассоциацию эти изменения не повлияли. В целом, результаты EMSA показали, что shTnsB связывает каждый повтор, присутствующий как в RE, так и в LE, независимым от груза способом.
Первоначально мы подготовили сетки крио-ЭМ и собрали данные, используя образец, содержащий shTnsB, а также RE и LE (т.е. две последовательности двухцепочечной ДНК, одна с последовательностью RE и одна с последовательностью LE без груза, дополнительный рисунок 2e). Эта выборка была неоднородной и страдала преимущественной ориентацией. Обработка этих данных привела к реконструкциям с низким разрешением недостаточного качества для построения модели атома (дополнительный рисунок 3). Однако можно было наблюдать два выступа, связанных с удлиненной плотностью, что свидетельствует о присутствии двух протомеров shTnsB, связанных с поверхностью двухцепочечной ДНК набора SR-LTR. Удлиненная плотность, приписываемая ДНК, изгибается таким же образом, как в недавно опубликованной структуре RE-связанного ecTnsB 9.1259 18 , что также подтверждает, что карта с низким разрешением соответствует комплексу перед транспозицией.
Чтобы стабилизировать комплекс shTnsB-ДНК, мы разработали олигонуклеотиды для восстановления STC, то есть состояния, представляющего посттранспозицию, а не претранспозицию, как описано выше (Методы, дополнительная таблица 1 и дополнительная рис. 4). Целевые последовательности для восстановленного комплекса были выбраны на основе природных последовательностей, фланкирующих сайт прикрепления в геноме S. hofmannii UTEX 2349. Эта стратегия аналогична той, которой следовали для получения структуры транспозосомы Р-элемента 20 , за исключением того, что в ней используется не симметричная ДНК STC, а природные асимметричные последовательности STC S. hofmannii UTEX 2349 , чтобы представить родной комплекс. ДНК STC состоит из двух перенесенных цепей (TS) и неперенесенных цепей (NTS), представляющих LTR(8)-SR(5) и SR(1)-LTR(6) двухцепочечную ДНК, связанную с сайтом прикрепления CAST (дополнительная рис. 4). Расстояние 5 п.н. между сайтами вставки LTR(8)-SR(5) и SR(1)-LTR(6) было выбрано, поскольку транспозиция CAST вызывает дупликацию 5 п.н. в сайте вставки 9.1259 21 (рис. 1а).
Воссозданная ДНК STC была смешана с очищенным белком, и сборка комплекса shTnsB-STC была проверена с помощью SEC-MALS (дополнительный рисунок 2b). Наблюдали два пика: пик с более низкой молекулярной массой содержал несвязанный белок и ДНК, использованные при восстановлении, в то время как пик с высокой молекулярной массой, который элюировался при 358,7 кДа, очень хорошо совпадал с ассоциацией ДНК STC (104,6 кДа) и 4 протомеров shTnsB (68 кДа), теоретическая молекулярная масса которого составляет 376,6 кДа. Эта сборка была подвергнута анализу отдельных частиц с помощью крио-ЭМ, в результате чего была получена карта с разрешением 2,46 Å, на которой мы построили атомную модель комплекса shTnsB-STC в его посткаталитическом состоянии (рис. 1а).
Общая структура комплекса shTnsB-STC
Полипептид shTnsB состоит из N-концевого домена, который можно разделить на два субдомена (NTD1 и NTD2), каталитического домена DDE (DDE), среднего домена (MD) , домен олигомеризации (OD) и С-концевой домен (CTD) (рис. 1b). Архитектура shTnsB напоминает архитектуру Mu-транспозазы (MuA) 22 , хотя идентичность последовательности ограничена каталитическим доменом DDE (дополнительный рисунок 1a). Мы собрали 4728 фильмов и установили координаты для 90,6 миллиона частиц, которые были уменьшены до 415 тысяч после 2D-классификации. Используя этот набор частиц, мы выполнили неравномерное уточнение, трехмерный анализ изменчивости и последующее гетерогенное уточнение в cryoSPARC 23 . Этот подход дал две карты при 2,5 и 2,8 Å. Дальнейшие этапы уточнения и трехмерного анализа изменчивости с использованием 260 тыс. частиц карты 2,5 Å позволили создать крио-ЭМ-карту с глобальным разрешением 2,46 Å, что позволило смоделировать комплекс shTnsB-STC (рис. 1c, дополнительные рис. 5–7, Дополнительная таблица 2 и методы). Однако большая гибкость ДНК поставила под угрозу визуализацию концов ДНК, содержащих SR (5) и SR (1), что не позволило определить возможные контакты с shTnsB (дополнительный рисунок 7, дополнительный фильм 1 и дополнительная таблица 1).
Структура комплекса shTnsB-STC напоминает вытянутую букву X с изогнутыми плечами разной длины. Более длинные плечи соответствуют концам транспозона, а короткие — изогнутой ДНК-мишени (рис. 1г, д, 2а). Четыре протомера shTnsB (shTnsB1-4) представляют собой переплетенную сборку на ДНК STC. Белок находится в двух разных конформациях, чтобы ассоциироваться с ДНК STC и строить комплекс (рис. 2b). shTnsB1 и 2 изображают вытянутую конформацию вдоль разветвленной структуры нуклеиновой кислоты, в которой мы не смогли обнаружить домены OD и CTD (рис. 1б, г). В обоих протомерах каждый из доменов NTD1 и NTD2 связан с LTR(8) и LTR(6), а каталитический домен DDE визуализируется в транс-положении на стыке ДНК-мишени и концов транспозона. Такое расположение каталитического домена в транс-положении является обычным при транспозиции ДНК, поскольку оно делает расщепление фосфодиэфира зависимым от сборки комплекса (рис. 1e, 2b). Со стороны ДНК-мишени сборка стабилизируется за счет взаимодействия остова с доменами MD и DDE протомеров shTnsB1 и 2 (R416, Q427, K29). 0, N428) (рис. 2б). Таким образом, shTnsB1 осуществляет распознавание ДНК на конце LTR(8), а его домен DDE катализирует атаку 3´OH в ответвлении LTR(6) на ДНК-мишень, способствуя реакции переноса цепи. Симметричное расположение ДНК STC наблюдается для shTnsB2, который катализирует перенос цепи LTR(8) на другую цепь ДНК-мишени (рис. 1а, 2б).
a На верхней панели показана схема S. hofmannii подвижный элемент с левым и правым концами (LE и RE) с каждой стороны ДНК-груза. Сильный цветовой тон показывает область каждого элемента, присутствующего в структуре. Нижняя панель не включает белковые фрагменты в структуру и показывает ДНК STC. ДНК окрашена по верхней схеме для LE и RE, а участок ДНК-мишени показан серым цветом. Желтые точки указывают сайт, где произошла прямая атака 3′-ОН на ДНК-мишень, чтобы способствовать реакции переноса цепи. b Подробное изображение ассоциации протомера shTnsB с ДНК STC. Две различные ассоциации наблюдаются для молекул shTnsB. Различные участки ДНК, взаимодействующие с белками, показаны соответствующим ярким цветовым оттенком. c Подробное изображение спиральных доменов NTD1 и 2, показывающее специфические взаимодействия с основаниями ДНК в области LTR(6). Маркировка нуклеотидов соответствует цветовому коду на рис. 1d, e (см. также рис. 1d и дополнительный рис. 8).
Изображение полного размера
Протомеры shTnsB3 и 4 расположены в другой конформации с небольшим количеством контактов с остовом ДНК (рис. 2б). Домены NTD1 и NTD2 этих протомеров не обнаружены в нашей структуре, что свидетельствует об их высокой подвижности. Кроме того, домен DDE вытягивается из ДНК, в то время как MD размещает области NTS с концов LTR (6) и LTR (8) после катализа. Структура выявила важную роль этих двух протомеров в ассоциации shTnsB1 и shTnsB2 с ДНК, поскольку в этой конформации они демонстрируют упорядоченный домен OD. Спираль между S502 и S522 не наблюдается в shTnsB1-2, но в конформации, принятой shTnsB3-4, спираль облегчает сборку комплекса STC за счет переплетения NTD2 shTnsB1 и shTnsB2 с доменом DDE противоположного протомера. в каждой ветви ДНК, тем самым связывая распознавание двух концов транспозона с каталитическими сайтами shTnsB1-2 (рис. 1d, 2b).
Распознавание shTnsB-ДНК
Распознавание LTR(6) и LTR(8) осуществляется спиральными доменами NTD1 и 2 субъединиц shTnsB1 и 2 (рис. 2c). Структура домена NTD1 обнаруживает спираль-поворот-спираль Myb/гомеодомен-подобную складку. Этот домен является членом суперсемейства гомеобоксных факторов транскрипции 24 . Прямые контакты оснований с помощью спирали, связывающей большую бороздку, в основном объясняют специфичное для последовательности распознавание. R77, который хорошо консервативен в гомологах цианобактерий и в Tn5053, но заменен лизином в MuA и аланином в ecTnsB (дополнительная рис. 1), образует полярные контакты с dG37, тогда как R81 связывается с основаниями последовательных нуклеотидов dG36 и dT37. на комплементарной цепи LTR (6) в LE (дополнительные рис. 7, 8 и дополнительная таблица 1). Другие остатки, взаимодействующие с ДНК (R58, R66, K84 и T78), обнаруживают полярные взаимодействия с остовом обеих цепей ДНК.
NTD1 соединен с NTD2 длинной петлей, которая проходит вдоль бороздки ДНК. Домен NTD2 функционально аналогичен второму ДНК-связывающему домену в других транспозазах. Однако он демонстрирует ограниченную консервативность по отношению к ближайшим гомологам shTnsB (дополнительный рисунок 1), а его структура напоминает парный домен, обнаруженный в генах парного ящика (Pax). Хорошо консервативный R99 в этой петле создает полярные взаимодействия с основаниями dT31 и dA32 и dA45, в то время как R106 также распознает основания dT30 и dT46 в разных цепях. Остальные ассоциации петли с ДНК включают остовные контакты. Домен NTD2 также связывается с ДНК в большой бороздке. Однако он отображает меньше конкретных контактов с базами. Только R158 и K154 связаны с dA48-dG49.и dG24 соответственно, тогда как остальные остатки связаны с остовом нуклеиновой кислоты. Эти взаимодействия аналогичны в доменах NTD1 и 2 на ветвях LTR (6) и LTR (8) комплекса белок-ДНК (дополнительная рис. 7).
Сборка комплекса белок-ДНК
Домены NTD2 субъединиц shTnsB1-2 соединены с доменами DDE длинной петлей. Переплетающаяся конформация этих протомеров с ДНК позволяет shTnsB1, который распознает ДНК в LTR(8), катализировать реакцию переноса цепи на конце LTR(6), и наоборот для shTnsB2 (рис. 1c, 2b). Этому перекрестному расположению каталитических доменов DDE способствует конформация молекул shTnsB3-4, которые стабилизируют сборку за счет интеркалирования длинной двудольной спирали, которая собирает домены NTD2 и DDE разных субъединиц. Соседний MD вмещает 5 ′ NTS (рис. 1d, рис. 3a–c). С-концевая спираль OD плотно прилегает между доменом DDE shTnsB2 и NTD2 shTnsB1 сетью контактов, сочетающих полярные и неполярные взаимодействия (рис. 3b), причем первая на конце (E360-K520 ), средний (R367-D512-Q508) и начальный (h372-S505) участки спирали. Другое сильное полярное взаимодействие наблюдается между боковыми цепями D514 в OD и R137 в NTD2. Короткая петля соединяет этот участок с N-концевой спиралью OD. Взаимодействие второй спирали OD с доменом DDE стабилизируется за счет неполярных взаимодействий, а группа остатков в MD shTnsB3 (R416, Q425, N428) вместе с R174 и R179в shTnsB2 образуют электроположительно заряженную область, стабилизирующую основания 5′-выступа (рис. 3c). Наконец, домены MD и DDE протомера shTnsB3 связаны с NTD1 shTnsB2, главным образом, группой неполярных взаимодействий. Интересно, что N-концевая β-цепь shTnsB2 встроена в качестве дополнительной цепи в антипараллельный β-лист MD (рис. 3d). Дополнительные полярные взаимодействия боковых цепей (E422-K169, N462-K102 и E54-h383) дополняют эту ассоциацию. В целом, описанные здесь ассоциации очень похожи в случае shTnsB4 с shTnsB1 и 2. Отметим, что взаимодействия между shTnsB3 и 4 в комплексе shTnsB-STC не наблюдается.
a Обзор архитектуры протомеров shTnsB и взаимодействий между двумя типами конформаций, наблюдаемых для белка на одной из ветвей ДНК STC (прозрачно). Цветные овалы обозначают увеличенные области на других панелях. b , c Детальное изображение взаимодействий OD shTnsB3 с доменами NTD2 и DDE shTnsB1 и 2 соответственно. д Увеличение области LTR(6), изображающей ассоциацию N-концевого β-листа NTD1 с MD shTnsB3.
Изображение полного размера
В совокупности эта сеть взаимодействий вдоль Х-образного комплекса предполагает, что различная конформация протомеров shTnsB1-2 и shTnsB3-4 благоприятствует транс-архитектуре, скорее всего, как стратегии связывания белок-ДНК. комплексная сборка с катализом ДДЕ.
Каталитический домен DDE
Транспозазы из нескольких суперсемейств обладают каталитическим доменом, содержащим триаду кислых аминокислот (DDE или DDD) 25,26 . Этот белковый модуль катализирует реакцию транспозиции, при которой элемент вырезается из донорского сайта и вставляется в геном или в мобильный генетический элемент. Мутации в этих каталитических остатках показали их критическую роль в транспозиции в доменах DDE транспозаз Tn5 и Tn10 27,28 . Интеграция элементов в новое место генома обычно приводит к короткой дупликации сайта-мишени из последовательностей хозяина (2–10 п.н.). В shTnsB мотив DDE состоит из двух остатков аспарагиновой кислоты (D205, D287) и остатка глутаминовой кислоты (E321), расположенных в консервативном ядре, образующем характерную РНКазу Н-подобную складку, объединяющую α-спирали и β-цепи (рис. 4а, дополнительный рис. 8е). Трехмерная структура триады ДДЭ образует каталитический карман, который связывается с ионами двухвалентного металла, которые помогают в различных нуклеофильных реакциях во время расщепления ДНК. Домены DDE четырех молекул shTnsB в сборке очень хорошо накладываются друг на друга (среднеквадратичное отклонение 0,59).Å среднеквадратичное значение более 160 Cα). Однако карман DDE демонстрирует два разных расположения в зависимости от конформации протомера shTnsB в комплексе STC (рис. 1). Карманы DDE shTnsB3 и 4 не контактируют с ДНК, и каталитические остатки этих субъединиц не расположены должным образом для реакции расщепления (рис. 4b). В случае shTnsB1 и 2 они обнаруживают каталитический карман DDE после завершения реакции переноса цепи. Можно визуализировать дополнительную плотность в кармане (рис. 4а), которую можно отнести к H 2 O или Mg 2+ . E321 находится на расстоянии 6 Å, так как структура захватывается в посткаталитическом состоянии. Расстояния дополнительной плотности до D205, D287 и фосфатов основной цепи позволяют предположить, что плотность может быть отнесена к молекуле H 2 O, образующейся после 3′-OH-атаки одной из цепей мобильного элемента на ДНК-мишень. (рис. 1а, 4а, дополнительный рис. 8).
a В обведенной области находится один из каталитических центров в комплексе shTnsB-STC. Увеличение области показывает конформацию каталитического центра после катализа. Предполагаемая молекула воды и расстояния до соседних каталитических остатков и ДНК показаны в Å. b Наложение доменов DDE двух конформаций, обнаруженных в комплексе белок-ДНК для shTnsB. c Суперпозиция доменов DDE shTnsB2 и 3 с ecTnsB (PDB: 7PIK) и основным доменом MuA (PDB: 1BCO). СКО для суперпозиции находятся в диапазоне от 0,7 до 1,2 Å для 160 и 250 Cα. d Нижняя панель показывает, что, несмотря на высокое структурное сходство доменов DDE, единственной субъединицей во всех этих структурах, которая демонстрирует компетентный каталитический карман, является shTnsB2, которая является единственной, связанной с ДНК.
Изображение полного размера
Несмотря на различия в последовательностях, новая структура комплекса распознавания концов транспозона Tn7 ecTnsB транспозазы 18 обеспечивает функциональный гомолог для сравнения. Домен DDE является наиболее консервативным модулем shTnsB, и поиск гомологов shTnsB DDE с использованием DALI 29 выявил домены ecTnsB и транспозазы фага Mu как наиболее близкие структурные гомологи вместе с различными вирусными интегразами (SRV, HIV- 1, ВТЛВ-1). Сравнение двух конформаций shTnsB с этими белками показало, что ядро домена очень хорошо накладывалось на другие интегразы (рис. 4c), но только каталитический карман shTnsB1-2 показал триаду кислых аминокислот на соответствующих расстояниях для катализа реакция переноса цепи (рис. 4d). В остальных случаях каталитические остатки не были правильно расположены. Примечательно, что протомеры ecTnsB в комплексе распознавания концов имеют конформацию, сходную с протомерами shTnsB3 и 4 в STC. Однако в последнем ДНК-связывающие домены этих двух субъединиц не визуализировались (рис. 1–2), что позволяет предположить, что высокая гибкость, наблюдаемая в дистальных отделах ДНК STC (дополнительный фильм 1), нарушает их связывание с SR(1) и SR(5) области LE и RE.
В совокупности структурное сравнение показывает, что домен DDE поддерживает очень хорошо сохранившуюся трехмерную структуру; однако сравнение различных структурных гомологов предполагает, что архитектура каталитического кармана транспозазы устроена правильно, как только она связывается с ДНК-мишенью.
Связывание ДНК транспозазами shTnsB, ecTnsB и MuA
Белок shTnsB проявляет гомологию с транспозазами ecTnsB и MuA. Однако, помимо домена DDE, между ними сохраняется несколько остатков (дополнительный рисунок 1). Эти различия особенно очевидны в ДНК-связывающих доменах, где многочисленные выравнивания последовательностей из нескольких канонических белков Tn7 TnsB и различных классов CAST-элементов показали существенное расхождение 18 . NTD1 shTnsB демонстрирует значительное трехмерное сходство с доменом HTH DBD1 в ecTnsB (RMSD 2,4 Å для 66 Cα) и доменом Iβ MuA (RMSD 2,8 Å для 62 Cα). Тем не менее, присутствие N-концевой β-цепи (N32-T36) исключает shTnsB, и эта область играет важную роль в построении сборки комплекса STC (рис. 3d). Эта структурная особенность не наблюдается в комплексе MuA-STC 22 , единственной доступной до сих пор структуре прокариотического комплекса транспозазы STC (рис. 5).
ДНК-связывающие домены трех различных транспозаз выровнены по ДНК (PDB: 8AA5, 7PIK и 4FCY соответственно). В случае MuA показаны одни и те же домены, связанные с разными повторами. Для shTnsB были построены только домены протомеров 1 и 2. В случае ecTnsB изображен ДНК-связывающий домен одной из субъединиц концевого комплекса транспозона.
Полноразмерное изображение
Интересно, что домен NTD2 shTnsB не проявляет трехмерного сходства с его аналогами в транспозазах ecTnsB и MuA (рис. 5). Вместо этого NTD2 хорошо накладывается на Pax6, фактор транскрипции, содержащий двудольный спаренный ДНК-связывающий домен (RMSD 2,8 Å для 60 Cα), который играет важную роль в развитии глаза, носа, поджелудочной железы и центральной нервной системы 30 . Удлиненный линкер, соединяющий NTD1 и 2, частично визуализируется в ecTnsB и не наблюдается в ДНК-связывающих доменах MuA (рис. 5). Этот линкер образует контакты малой бороздки, а карбоксиконцевая единица спираль-поворот-спираль образует контакты основания в большой бороздке, как это наблюдается в комплексе shTnsB-STC (рис. 5).
Сравнение позволяет предположить, что белки TnsB имеют общий общий способ связывания ДНК, включая два домена, которые распознают разные SR и LTR в сборке. Однако домены, участвующие в связывании, претерпевают различные структурные вариации для осуществления специфических взаимодействий белок-ДНК для каждой транспозазы. Это разнообразие может быть необходимо для размещения последовательностей большого количества Tn7-подобных элементов, предотвращая распознавание транспозазами последовательностей ДНК, принадлежащих другим транспозонам 31 .
Комплексы STC
Структура концевого комплекса транспозона ecTnsB предоставила информацию о распознавании концов транспозона 18 , но до сих пор нет структурной информации об их комплексе STC. Конформация субъединиц ecTnsB в концевом комплексе транспозона напоминает расположение, принятое доменом DDE протомеров shTnsB3 и 4 в комплексе shTnsB-STC. Однако положения доменов MD и OD ориентированы по-разному (рис. 4c). Тем не менее, эта конформация сильно отличается от архитектуры протомеров 1 и 2 shTnsB, которые участвуют в катализе реакции переноса цепи (рис. 2b). Конформация этих субъединиц больше похожа на аналоги в комплексе MuA-STC (рис. 6а).
Рис. 6: Анализ комплексов переноса цепи shTnsB и MuA. a Комплексы shTnsB-STC и MuA-STC показаны в той же цветовой схеме, что и на рис. 1, за исключением ДНК, окрашенной в белый цвет. b Обведенные кружками области на обоих комплексах сравниваются, чтобы показать ключевые различия в сборке комплексов STC.
Полноразмерное изображение
Хотя последовательность shTnsB отдаленно связана с транспозазой фага MuA (дополнительная рис. 1), общая ассоциация комплекса MuA-STC имеет сходство с белком CAST (рис. 6a). Однако сборка также имеет важные отличия. Одно из основных отличий заключается в том, что в комплексе MuA-STC ДНК-связывающие домены субъединиц, участвующих в сборке (MuA 3 и 4), связаны с соответствующими повторами в LE и RE (рис. 6а), тогда как в шТнсБ-СТЦ их нельзя было смоделировать, что свидетельствует об их высокой гибкости. Это мнение подтверждается высокой изменчивостью, наблюдаемой во внешней части Х-образной структуры ДНК (дополнительный фильм 1). Тот факт, что эти области структуры MuA-STC 22 участвуют в контактах кристаллов, что может снижать гибкость, облегчая тем самым связывание доменов Iβ и Iγ с повторами на LE и RE концах.
Любопытно, что ассоциация уникальной N-концевой β-цепи NTD1 в shTnsB не наблюдается в MuA (рис. 6а, б, верхние панели), поскольку этот элемент вторичной структуры отсутствует ни в домене Iβ MuA, ни в DBD1 ecTnsB. Это наблюдение предполагает, что в комплексе ecTnsB-STC эта ассоциация не может происходить. Другое важное отличие возникает из-за сборки комплекса, в котором домен OD в shTnsB интеркалирует с множественными взаимодействиями между NTD2 и каталитическим доменом DDE, создавая ассоциацию между белками shTnsB. В случае MuA ассоциация между четырьмя протомерами в этой области комплекса STC отличается, демонстрируя значительно меньше взаимодействий между разными доменами (рис. 6b). Кроме того, в протомерах 1 и 2 комплекса shTnsB-STC домены OD не наблюдаются, тогда как в случае MuA спирали в домене IIIα перекрещиваются внутри V-образного участка ДНК-мишени (рис. 6а). ). Нельзя было не учитывать возможность того, что взаимодействие между этими спиралями в структуре MuA происходит за счет кристаллических контактов.
Эффект связывания ДНК при транспозиции
Чтобы подтвердить взаимодействие shTnsB:ДНК в STC, мы протестировали транспозиционную активность ряда мутантов с заменой, в которых консервативные аминокислоты, демонстрирующие полярные взаимодействия с ДНК, были заменены аланином (рис. 7, дополнительный рис. 1). Мы мутировали R77, R81 (оба присутствуют в NTD1), R99 (присутствуют в линкере между NTD1 и NTD2), R158 (присутствуют в NTD2), R188, R223, K290 и R380 (присутствуют в домене DDE) (рис. 7b). . Мутанты R77A, R81A и R158A продемонстрировали резкое влияние на активность транспозиции «на-мишень», предполагая, что shTnsB больше не может распознавать повторы в RE и LE (рис. 7b). Примечательно, что нуклеотиды, участвующие в этом взаимодействии (дополнительная рис. 7), сохраняются между различными повторами, что еще раз подтверждает важность этого взаимодействия в распознавании ДНК донора 21 . Что касается R99, хотя он и присутствует в неупорядоченной области shTnsB, он тесно взаимодействует с фосфатным остовом и основаниями LTR. Эти нуклеотиды также консервативны в наборе повторов RE/LE 21 . Однако эффект его замены был переменным: повторы демонстрировали либо более высокую, либо более низкую активность, чем дикий тип. В случае R223A и R380A это также повлияло на активность. Эти остатки, по-видимому, участвуют в стабилизации состояния после транспозиции. R223 взаимодействует с центральными нуклеотидами последовательности 5 bp между никами, образованными транспозицией, тогда как R380 взаимодействует с нерасщепленной одноцепочечной ДНК транспозона, расположенной вне комплекса. Наконец, несмотря на свою консервативность и близость к сайту прикрепления, мутанты R188A и K290A не отменял целевую транспозицию. Наши результаты подтверждают функциональную важность этих остатков, участвующих в распознавании LTR и стабилизации STC, и указывают на возможные сайты для улучшения свойств транспозазы для целей инженерии генома.
a Транспозицию проводили путем трансформации бактерий плазмидой, содержащей сайт-мишень, показанный зеленым цветом, распознаваемым Cas12k:sgRNA (pTarget), плазмидой, содержащей донорную ДНК с последовательностями CAST RE и LE, показанными красным ( pDonor) и плазмиду, содержащую кодирующие последовательности sgRNA, Cas12k, TnsC, TniQ и TnsB (pHelper). Интеграцию донорной последовательности в pTarget количественно определяли с помощью количественной ПЦР с использованием набора праймеров, амплифицирующих область pTarget, в которой не происходит транспозиции (PF 9).1331-мишень и PR -мишень ) и набор праймеров, которые амплифицируют область между последовательностями LE-мишени и донора (продукт PF и продукт PR ). b Положение выбранных аминокислот в STC, подлежащих мутации и тестированию на транспозиционную активность. c Кратность изменения активности, измеренная с помощью количественной ПЦР для каждого мутанта TnsB. Для каждого мутанта проводили две или три биологические повторности. Отдельные точки данных, представляющие все технические повторы, обозначены точками. Столбцы представляют собой среднее значение кратности изменения активности, а усы, расположенные вокруг него, представляют собой стандартное отклонение. Активность дикого типа представлена пунктирной линией. Данные предоставляются в виде файла исходных данных.
Изображение в натуральную величину
Обсуждение
В прошлом году мы стали свидетелями значительного увеличения структурных знаний систем CAST. Доступна структурная информация TnsC, TniQ и Cas12k типа V-K 32,33,34 , а также каскадного комплекса типа I-F, связанного с TniQ 35,36 . Однако, несмотря на эти достижения, нам необходимо точное понимание последовательности взаимодействий и промежуточных звеньев, ведущих к точной вставке ДНК-груза, чтобы решить возможную разработку систем CAST. Это отсутствие полного понимания того, как транспозиция осуществляется с помощью различных CAST, препятствует их повторному использованию для точной вставки ДНК в инструменты редактирования генов. Тип В-К мог представлять собой перспективную систему для развития этих средств за счет уменьшенного количества и размеров ее компонентов. Однако для этой цели необходимо решить проблему образования коинтегратов (рис. 1а).
Концевой комплекс транспозона shTnsB с низким разрешением (дополнительный рисунок 3) предполагает, что распознавание конца транспозона аналогично ecTnsB 18 ; однако высокая гетерогенность и гибкость сборки не позволили провести детальную молекулярную характеристику распознавания ДНК. Это согласуется с экспериментами в системе ecTn7, которые показали, что пре-транспозиционный комплекс менее стабилен, чем пост-транспозиционный комплекс, и что защита транспозонов меняется между пре- и посткаталитической стадиями 17,37 . Кроме того, эксперименты EMSA предполагают, что shTnsB может взаимодействовать с RE и LE в прекаталитическом состоянии в отсутствие ДНК-мишени (дополнительный рисунок 2). Таким образом, взаимодействие shTnsB с концами транспозона, по-видимому, не зависит от присутствия других компонентов системы CAST типа V-K. Более высокая стабильность посткаталитических комплексов привела нас к сборке shTnsB-STC, которую мы смогли определить с высоким разрешением. Этот комплекс представляет собой стадию после завершения реакции переэтерификации с ДНК-мишенью, обнаруживающую ее трехмерную сборку. Общая структура напоминает архитектуру транспозосомы 9 фага MuA.1259 22 . Тем не менее, можно наблюдать ключевые различия между сборками, особенно в домене NTD2 (рис. 6). Одно сходство между shTnsB и MuA, которое отличает их от системы Tn7, заключается в том, что типы V-K и Mu используют репликативную транспозицию, которая требует расщепления только одной цепи ДНК, в то время как Tn7 выполняет реакцию вырезания и вставки, включающую расщепление неперенесенной цепи TnsA. Следовательно, структурное сходство общего комплекса STC между shTnsB и MuA будет поддерживаться этими механистическими сходствами. Тем не менее, представляется затруднительным установить аналогии возможной комплексной сборки STC ecTnsB 9.1259 18 с использованием структуры shTnsB-STC, так как мы ожидаем, что ecTnsB-STC также должен вмещать ecTnsA 10 . То же самое может быть экстраполировано для CAST типов I-F и I-B, которые включают в свой состав TnsA.
Посткаталитическая структура shTnsB-STC выявила очень мало оснований-специфических контактов белок-ДНК (дополнительная рис. 7), как это также наблюдалось в прекаталитическом комплексе ecTnsB 18 . Домен NTD1 в shTnsB частично имеет такую же архитектуру, что и его аналоги в ecTnsB и MuA, в то время как NTD2 уникален в том смысле, что он имеет структурную гомологию с фактором транскрипции Pax6 (рис. 5). N-концевая β-цепь, связывающая распознавание в одном из плеч транспозона с каталитическим центром на противоположной стороне и наоборот, является сингулярной особенностью shTnsB, однако еще предстоит определить, оказывает ли эта особенность некоторую аллостерическую регуляцию в каталитическом сайте DDE. . Две субъединицы shTnsB, участвующие в реакции, демонстрируют кислотную триаду в посткаталитическом состоянии. Атака 3′-ОН цепей транспозона на 5′ ДНК-мишени оставляет несвязанные основания (dT62 и dA63) в каждой цепи и молекулу воды, связанную с остатками аспарагиновой кислоты и остовом ДНК (дополнительная рис. 7) . На основании структуры концевого комплекса shTnsB-STC и концевого комплекса транспозона ecTnsB можно сделать вывод, что активный карман белков TnsB не приспособлен для катализа до связывания ДНК-мишени (рис. 4d). Скорее всего, этот механизм сочетается с транс-сборкой STC, чтобы предотвратить неспецифическое расщепление.
ShTnsB вставляет транспозон со смещением направленности 16 . Хотя концы LTR8-SR5 и LTR6-SR1 почти симметричны, длина и последовательности с правого и левого концов различаются (дополнительный рисунок 2f), что позволяет предположить, что эти различия в сочетании с необходимостью PAM для определения вставки сайте, может повлиять на направленность грузовой ДНК. Это мнение подтверждается наблюдением, что расстояние между PAM и местом прикрепления транспозона показывает очень узкое распределение 16 . Однако механизм, управляющий интеграцией ДНК-груза, до конца не ясен. Были предложены две альтернативные модели 32,34 . В обоих случаях сборка филамента shTnsC будет работать как молекулярная линейка, направляющая shTnsB к месту прикрепления. Действительно, С-концевая область shTnsB взаимодействует с shTnsC 32,34 , и взаимодействие shTnsB-shTnsC приводит к разборке филамента. Однако одна модель предполагает, что филамент shTnsC растет из shCas12k, а ассоциация головки филамента с shTniQ указывает на место вставки shTnsB и груз ДНК 32 . Вторая модель предполагает, что филамент shTnsC растет в противоположном направлении, и комплекс shCas12k-shTniQ останавливает рост, рекрутируя shTnsB с грузом ДНК, таким образом продолжая интеграцию 34 . Обе модели ставят дополнительные вопросы относительно взаимодействия shTnsB-shTnsC. Как показано в структуре shTnsB-STC, не все протомеры имеют одинаковую конформацию, и неясно, все ли протомеры shTnsB, каталитически активные или неактивные протомеры, взаимодействуют с shTnsC (рис. 1d, e), поскольку в обоих случаях C -конец в структуре не визуализируется. Однако, по-видимому, он топологически доступен для дополнительных взаимодействий. В свете современных биохимических и структурных данных 32,34 , мы хотели бы предложить альтернативную гипотетическую рабочую модель (рис. 8), которая считает, что shTnsC и shTnsB будут связаны до того, как shTnsC взаимодействует с ДНК, чтобы избежать неспецифических взаимодействий с геномом, которые могут быть токсичными для клетка. Таким образом, образование филаментов shTnsC будет динамичным, так как присутствие shTnsB вызовет диссоциацию сборки в отсутствие shCas12k и shTniQ. В этом сценарии ассоциация shCas12k с ДНК-мишенью и shTniQ стабилизирует сборку комплексов shTnsC-shTsnB в месте вставки, а затем запускает интеграцию груза ДНК. Необходим дальнейший анализ ассоциации shTnsB-shTnsC, чтобы полностью понять интеграцию ДНК-груза в CAST типа V-K и подтвердить любую из этих рабочих моделей.
ShTnsC связывается с ДНК, образуя филаменты, которые деполимеризуются с помощью shTnsB (слева). Учитывая взаимодействия shTnsC с shTnsB и shCas12k-shTniQ, возникают две возможности (штриховые квадраты). Во-первых, shTnsC, связанный с shTnsB, может рекрутироваться с помощью нацеливающего комплекса, включающего shCas12k и shTniQ (верхний пунктирный квадрат). Во-вторых, shTnsC может сначала связываться с нацеливающим комплексом, а затем рекрутировать shTnsB (нижний пунктирный квадрат). Обе возможности продуцируют комплекс перед транспозицией, активирующий активность интегразы shTnsB и приводящий к вставке транспозона.
Полноразмерное изображение
Отсутствие TnsA в типе V-K приводит к дополнительным вопросам относительно промежуточного разрешения интеграции, поскольку нерасщепленные 5′-концы транспозона приводят к продуктам коинтеграции транспозиции 15,16 . Транспозоны, полагающиеся на транспозазы DDE, имеют разные механизмы для решения этой проблемы 26,38,39 . Например, Tn5053 содержит резольвазу TniR, которая использует сайты res , также присутствующие в транспозоне, для разрешения коинтеграта 9.1259 40,41 , в то время как другие транспозазы генерируют шпильки для вырезания оставшейся ДНК 42 . Однако отсутствие домена вставки, который стабилизирует шпильку в каталитическом домене DDE shTnsB, не поддерживает эту возможность предотвращения коинтеграций. Кроме того, репликация ДНК и гомологичная рекомбинация могут разрешить коинтеграт в случае репликативной транспозиции 26 .
С другой стороны, наличие специфичной для хозяина резольвазы подтверждается тем фактом, что при транспозиции в E. coli , 20% продуктов внедрения являются коинтегратами 16 . Действительно, набор генов системы V-K (TnsB, TnsC и TniQ) сходен с набором генов Tn5053, который содержит TniA, TniB (гомологи TnsB и TnsC) и TniQ 16 , и можно предположить, что резольваза не кодируется в Локус транспозона VK, но все еще присутствующий в естественном хозяине, может иметь ту же активность, что и резольваза TniR. Необходимо провести дальнейшие исследования на S. hofmannii или других родственных цианобактериях, прежде чем мы сможем полностью понять РНК-управляемую транспозицию в типе V-K. Решение этих вопросов является ключом к успешному созданию новых комплексов CAST, способных интегрировать большие ДНК-грузы в геномы эукариот.
Наша структура обеспечивает механистическое понимание того, как CAST shTnsB распознает последовательности ДНК RE и LE транспозона в STC. Структура также показывает конформационную гимнастику белка shTnsB для сборки с ДНК и дает подробное представление об активном центре после катализа. Полное выяснение механизмов сборки shTnsB с остальными компонентами CAST потребуется, чтобы использовать системы CAST для РНК-управляемой геномной инженерии.
Методы
Получение и очистка белков
Кодирующую последовательность (IDT) Scytonema hofmannii (UTEX 2349) tnsB (IDT) клонировали с C -концевой гексагистидиновой (His)-меткой в вектор pET-21 (дополнительная таблица). Белки экспрессировали в клетках E. coli BL21 pRARE. Культуры E. coli выращивали при 37 °C в среде Terrific Broth (TB) с ампициллином в концентрации 100 мг/л до ОП 600 , равной 0,6. Сверхэкспрессию белков индуцировали 250 мМ ИПТГ в течение 16–18 ч при 18°С. Клетки собирали центрифугированием, мгновенно замораживали в жидком азоте и хранили при -80°С. Затем осадки ресуспендировали в лизирующем буфере (50 мМ HEPES pH 7,5, 500 мМ NaCl, 5 мМ MgCl 9). 1331 2 , 1 мМ TCEP, 1 таблетка полного коктейля ингибиторов без ЭДТА (Roche) на 50 мл, 50 ЕД/мл бензоназы, 2 мг на 50 мл лизоцима). Ресуспендированные осадки перемешивали в течение 1 ч при 4°С. Лизис завершали обработкой ультразвуком, а лизат центрифугировали при 10 000 × г в течение 45 минут для разделения растворимой и нерастворимой фракций. Растворимую фракцию загружали в 5 мл колонку HisTrap FF Crude (Cytiva), уравновешенную в буфере IMAC-A (50 мМ HEPES, pH 7,5, 500 мМ NaCl, 1 мМ TCEP, 10 мМ имидазол, 1 таблетка полного коктейля ингибиторов, не содержащего ЭДТА ( Roche) на 500 мл), а связанные белки элюировали ступенчатым повышением концентрации имидазола буфером IMAC-B (50 мМ HEPES pH 7,5, 500 мМ NaCl, 1 мМ TCEP, 500 мМ имидазол, 1 таблетка коктейля «Полный ингибитор»). без ЭДТА (Roche) на 500 мл). Обогащенные белковые фракции объединяли, а концентрацию NaCl снижали до 200 мМ с использованием буфера для разбавления (50 мМ HEPES, pH 7,5, 50 мМ NaCl, 1 мМ TCEP). Образец наносили на колонку HiTrap Heparin HP (GE Healthcare), уравновешенную буфером Heparin-A (50 мМ HEPES, pH 7,5, 200 мМ NaCl, 1 мМ TCEP). Белок элюировали линейным градиентом 0-100% буфера Heparin-B (50 мМ HEPES pH 7,5, 1 мМ NaCl, 1 мМ TCEP). Фракции, содержащие белок, объединяли и смешивали с TEV-протеазой для расщепления гистидиновой метки. Затем белок концентрировали и дополнительно очищали методом эксклюзионной хроматографии (SEC) с использованием колонки HiLoad 16/600 Superdex 200 (Cytiva), уравновешенной буфером SEC (50 мМ HEPES, pH 7,5, 500 мМ NaCl, 1 мМ TCEP). Фракции, содержащие чистый белок, объединяли, концентрировали до 9мг/мл, мгновенно замороженные в жидком азоте и хранящиеся при температуре -80 °C.
Производство ДНК
Последовательности ДНК, соответствующие отдельным повторам, двойным повторам и одноцепочечной одноцепочечной ДНК, были приобретены у IDT. Молекулы ДНК, содержащие три или более повторов, были получены с помощью ПЦР с донорной ДНК, клонированной в pUC57 (Genewiz) в качестве матрицы (дополнительная таблица 1). Последовательности ДНК были выбраны из природного транспозона для CAST типа V-K в S. hofmannii , описанном в 4,7,21. Полную ДНК донора амплифицировали с использованием праймеров 5′-tgctacgtctctacgtgtacagtgactaat-3′ и 5′ ccacaaaaggcgtagtgtacagtgacaaat-3′, RE амплифицировали с использованием праймеров 5′-tgctacgtctctacgtgtacagtgactaat-3′ и 5′-atacaaaagcgacagtcaatttgtcattatgaaa -attatattgatgacatttaatttgtcatcaattaattaag-3′ и 5′-ccacaaaaggcgtagtgtacagtgacaaat-3′. Амплифицированные последовательности очищали с использованием набора QIAquick PCR Purification Kit (Qiagen) и элюировали водой, не содержащей нуклеаз.
Анализ сдвига электромобильности (EMSA)
ДНК (донорскую, RE, LE или LTR(6)-SR(1)) смешивали с shTnsB в буфере, содержащем 50 мМ HEPES pH 7,5, 300 мМ NaCl, 5 мМ MgCl 2 и 1 мМ TCEP. Концентрации ДНК и shTnsB указаны на дополнительных рисунках 3c – e. Образцы инкубировали при 37 °C в течение 30 мин и загружали в гели для замедления ДНК 6% (Thermo Fisher Scientific). ДНК выявляли окрашиванием геля GelRed (VWR). Затем тот же гель окрашивали InstantBlue (Life Technologies) для обнаружения присутствия shTnsB и сравнения сдвигов полос.
Крио-ЭМ пробоподготовка
Прекаталитический комплекс готовили с использованием 1 мкМ РЭ, 1 мкМ ЛЕ и 8 мкМ шТнсБ в 300 мкл буфера, содержащего 50 мМ HEPES рН 7,5, 300 мМ NaCl, 5 мМ MgCl. 2 и 1 мМ TCEP. Образец инкубировали в течение 30 мин при 37°С и концентрировали до 50 мкл в фильтрах 10 кДа. Восстановление STC показано на дополнительном рисунке 4 и проводилось следующим образом. Перенесенные и неперенесенные нити (TS и NTS) из RE (олигонуклеотиды RE_Target и RE_polyA в дополнительной таблице 1) и LE (олигонуклеотиды LE_Target и LE_polyA в дополнительной таблице 1) смешивали по отдельности, инкубировали при 95 °C в течение 5 мин и оставили на льду на 10 мин. Впоследствии комплементарная одноцепочечная ДНК-мишень была добавлена к двухцепочечной ДНК TS-NTS из RE и LE (олигонуклеотиды Target_1 и Target_2 в дополнительной таблице 1 соответственно). ДцДНК TS-NTS из RE и LE инкубировали отдельно при 70 ° C в течение 5 минут и оставляли на льду на 10 минут. Затем их перемешивали и инкубировали при комнатной температуре в течение 10 мин. Наконец, 50 мМ HEPES pH 7,5, 300 мМ NaCl, 5 мМ MgCl 2 и 1 мМ TCEP добавляли к 10 мкМ ДНК STC ДНК перед добавлением 40 мкМ shTnsB. Образец инкубировали в течение 30 мин при 37°С.
Сетки для обоих образцов (прекаталитический комплекс и STC) были приготовлены путем нанесения 3 мкл свежеприготовленного комплекса (разведение 1:2) на сетки с отверстиями UltraAuFoil 300 меш R1,2/1,3 (Quantifoil), подвергнутые тлеющему разряду в течение 60 с при 10 мА (Leica EM ACE200) и замораживание погружением в жидкий этан (предварительно охлажденный жидким азотом) с использованием Vitrobot Mark IV (FEI, Thermo Fisher Scientific — время блоттинга 3 с, влажность 100%, 4 °C) .
Крио-ЭМ обработка данных
Сбор данных проводили с помощью EPU 2.12.1 (ThermoFisher). Обработку данных проводили с помощью программы cryoSPARC версии 3. 3.2 23 . Выполняли коррекцию движения заплаты и оценку CTF заплаты. Частицы 9646 K были первоначально отобраны из 4574 микрофотографий с использованием устройства для сбора капель с минимальным и максимальным диаметром 50 и 200 Å соответственно, а затем извлечены и отсортированы с помощью двух раундов 2D-классификации. Выбранные 2D-классы содержали 415 K частиц, извлеченных с использованием блока размером 416 × 416 пикселей. Трехмерная реконструкция ab initio использовалась в качестве начального объема для неравномерного уточнения, а полученные частицы были проанализированы с помощью трехмерного анализа изменчивости 43 (дополнительный рисунок 5), который выявил непрерывную конформационную изменчивость дистальных концов ДНК и близлежащих доменов shTnsB (N-концевая часть и CTD цепей A и B). Два наиболее пространственно отличающихся полученных объема использовались в качестве эталонов для гетерогенного уточнения и были дополнительно уточнены с помощью неоднородного (NU) уточнения до глобального разрешения 2,47 Å (частицы 269 K) и 2,81 Å (частицы 155 K) на основе золотой стандарт корреляции оболочки Фурье (GSFSC) 0,143 критерий отсечки (дополнительный рисунок 6). Параметры расфокусировки для каждой частицы и параметры CTF для каждой группы были уточнены во время уточнения NU. При визуальном осмотре плотность карты 2,47 Å оказалась значительно более непрерывной. Эта карта была дополнительно улучшена на дистальных концах с помощью еще одной итерации трехмерного анализа изменчивости частиц 43 и использование наиболее пространственно различных объемов в качестве эталонов для еще одного раунда неоднородного уточнения. Большинство частиц попали в один класс (208 K), который был окончательно уточнен с помощью уточнения NU до карты 2,46 Å. Анализ 3DFSC показал сферичность 0,928 и диапазон углового разрешения 2,4–3,1 Å (дополнительный рисунок 6). Анализ локального разрешения был выполнен с помощью MonoRes 44 с использованием обертки cryoSPARC, который показал диапазон ~ 2–10 Å, при этом большая часть составляла около 2,5 Å, а дистальная часть — 4–8 Å. Построение модели выполнялось на картах, прошедших постобработку с помощью DeepEMhancer 0,13 45 . В качестве входных данных использовались полукарты, в то время как нормализация и маскирование/шумоподавление входных объемов выполнялись автоматически в режиме tightTarget, что давало карты со значительно улучшенной интерпретируемостью. Окончательная модель, построенная на картах DeepEMhancer, была окончательно проверена с использованием экспериментальных карт.
Построение и уточнение атомной модели
Исходная модель была создана на основе структуры MuA (PDB: 4FCY) с использованием автоматизированного сервера моделирования гомологии белковой структуры SWISS-MODEL 9.1259 46 . Это был жесткий корпус, встроенный в карту с помощью ChimeraX 47 . Подгонка модели была проверена и дополнительно подогнана в COOT 48 , а менее упорядоченные N- и C-концевые области были удалены. Модель Alphafold 49 мономера shTnsB была впоследствии использована для включения некоторых из этих областей в карту. Далее модель была перестроена de novo в COOT и уточнена с использованием phenix. real_space_refine 50 , в то время как сервер гибкой настройки Namdinator молекулярной динамики использовался для ускорения и улучшения модели для сопоставления соответствия и для уточнения модели как на ранней, так и на поздней стадии. этапы доработки 51 .
Анализ транспозиции и мутагенез in vivo
Активность транспозиции CAST определяли с использованием векторов pHelper, pTarget и pDonor, полученных от Addgene (кат. № 127922, 127926 и 127924). Мутанты shTnsB (рис. 7) были созданы в векторе pHelper с использованием набора для клонирования In-Fusion от Takara и праймеров, содержащих желаемые точечные мутации. Анализы in vivo проводили в соответствии с Saito et al. 52 . Вкратце, по 12 нг pHelper, pTarget и pDonor были преобразованы в 30 мкл TransforMax EC100 pir+ Electrocompetent 9.1267 E. coli (Люциген). Клетки восстанавливали в течение 1 ч и высевали на среду LB, содержащую ампициллин, канамицин и хлорамфеникол. Клетки соскребали через 24 часа после посева, лизировали в 15 мкл буфера для лизиса колоний (TE с 0,1% Triton X-100), кипятили в течение 5 минут, разбавляли 30 мкл воды и вращали при 4000× г в течение 10 минут для осадка. обломки. Супернатант использовали для последующего анализа.
Анализ активности кПЦР
Частоту вставок относительно концентрации целевой плазмиды определяли с помощью кПЦР (TaqMan Fast Advanced Master Mix, Applied Biosytems) с набором праймеров/зонда (5′-6-FAM, Int ZEN и набор праймеров/зондов (5′-6-FAM, Int ZEN, 3′-Iowa Black, IDT), соответствующих продукту транспозиции (прямой праймер: 5′-ggttgagaagtcatttaataaggccactgttaaacg-3′, обратный праймер: 5′-aacgctgatgggtcacgacg- 3′, зонд: 5′-ctgtcgtcggtgacagattaatgtcattgtgac-3′) (рис. 7). Каждая реакция (11 мкл) содержала 2 мкл экстрагированных нуклеиновых кислот, 900 нМ прямого праймера, 900 нМ обратного праймера и 250 нМ зонда. Флуоресценцию измеряли с использованием прибора LightCycler 480 (Roche). Данные анализировали методом 2 –∆∆Ct с нормализацией по pTarget Ct.
Сводка отчета
Дополнительная информация о дизайне исследования доступна в Сводке отчета об исследовании природы, связанной с этой статьей.
Доступность данных
Реагенты, полученные в ходе этого исследования, предоставляются по запросу с заполненным Соглашением о передаче материалов. Атомные координаты и крио-ЭМ карты, включенные в это исследование, были депонированы в банке данных белков и банке данных электронной микроскопии под кодами доступа: PDB 8AA5, EMD-1529.4 и EMD-15344. Исходные данные qPCR и несрезанные гели предоставляются с этой статьей. Исходные данные приводятся вместе с настоящей статьей.
Список литературы
Макарова К.С. и др. Эволюционная классификация систем CRISPR-Cas: взрыв класса 2 и производные варианты. Нац. Преподобный Микробиолог. 18 , 67–83 (2020).
Артикул КАС пабмед Google Scholar
Дудна, Дж. А. Перспективы и проблемы терапевтического редактирования генома. Природа 578 , 229–236 (2020).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Фор, Г. и др. CRISPR-Cas в мобильных генетических элементах: контрзащита и не только. Нац. Преподобный Микробиолог. 17 , 513–525 (2019).
Артикул КАС пабмед Google Scholar
Петерс Дж. Э., Макарова К. С., Шмаков С., Кунин Е. В. Рекрутирование систем CRISPR-Cas Tn7-подобными транспозонами. Проц. Натл акад. науч. США 114 , E7358–E7366 (2017).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Кунин Е.В., Макарова К.С. Происхождение и эволюция систем CRISPR-Cas. Филос. Транс. Р. Соц. Лонд. Б биол. науч. 374 , 20180087 (2019).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Рыбарски Дж. Р., Ху К., Хилл А. М., Уилке К. О. и Финкельштейн И. Дж. Метагеномное открытие CRISPR-ассоциированных транспозонов. Проц. Натл акад. науч. США , https://doi.org/10.1073/pnas.2112279118 (2021).
Шмаков С. и др. Разнообразие и эволюция систем CRISPR-Cas класса 2. Нац. Преподобный Микробиолог. 15 , 169–182 (2017).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Klompe, S.E., Vo, PLH, Halpin-Healy, T.S. & Sternberg, S.H. Системы CRISPR-Cas, кодируемые транспозонами, обеспечивают прямую интеграцию ДНК под управлением РНК. Природа 571 , 219–225 (2019).
Hickman, A.B. et al. Неожиданное структурное разнообразие в рекомбинации ДНК: рестрикционная эндонуклеазная связь. Мол. Ячейка 5 , 1025–1034 (2000).
Артикул КАС пабмед Google Scholar
Чой, К.Ю., Ли, Ю., Сарновский, Р. и Крейг, Н.Л. Прямое взаимодействие между субъединицами TnsA и TnsB контролирует гетеромерную транспозазу Tn7. Проц. Натл акад. науч. США 110 , E2038–E2045 (2013 г.).
Stellwagen, AE & Craig, NL. Анализ мутантов с усилением функции АТФ-зависимого регулятора транспозиции Tn7. Дж. Мол. биол. 305 , 633–642 (2001).
Артикул КАС пабмед Google Scholar
Сарновский Р. Дж., Мэй Э. У. и Крейг Н. Л. Транспозаза Tn7 представляет собой гетеромерный комплекс, в котором активность разрыва и соединения ДНК распределяется между различными генными продуктами. EMBO J. 15 , 6348–6361 (1996).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Питерс, Дж. Э. Tn7. Микробиолог. Спектр. 2 , 1–20 (2014).
Кудувалли, П. Н., Рао, Дж. Э. и Крейг, Н. Л. Структура ДНК-мишени играет решающую роль в транспозиции Tn7. EMBO J. 20 , 924–932 (2001).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Райс, П. А., Крейг, Н. Л. и Дайда, Ф. Комментарий к «Вставка ДНК под управлением РНК с CRISPR-ассоциированными транспозазами». Наука , https://doi.org/10.1126/science.abb2022 (2020).
Стрекер, Дж., Ладха, А., Макарова, К.С., Кунин, Е.В. и Чжан, Ф. Ответ на комментарий к «РНК-управляемая вставка ДНК с CRISPR-ассоциированными транспозазами». Наука , https://doi.org/10.1126/science.abb2920 (2020).
Холдер, Дж. В. и Крейг, Н. Л. Архитектура посттранспозиционного комплекса Tn7: сложная нуклеопротеиновая структура. Дж. Мол. биол. 401 , 167–181 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Kaczmarska, Z. et al. Структурная основа распознавания концов транспозонов объясняет центральные особенности систем транспозиции Tn7. Мол. Сотовый , https://doi.org/10.1016/j.molcel.2022.05.005 (2022).
Дэвис Д. Р., Горышин И. Ю., Резникофф В. С. и Реймент И. Трехмерная структура промежуточного продукта транспозиции синаптического комплекса Tn5. Наука 289 , 77–85 (2000).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед Google Scholar
Ghanim, G.E., Kellogg, E.H., Nogales, E. & Rio, D.C. Структура комплекса транспозаза-ДНК P-элемента выявляет необычные структуры ДНК и контакты GTP-ДНК. Нац. Структура Мол. биол. 26 , 1013–1022 (2019).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Strecker, J. et al. Вставка ДНК под управлением РНК с CRISPR-ассоциированными транспозазами. Наука 364 , 48–53 (2019).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Montaño, S.P., Pigli, Y.Z. & Rice, P.A. Структура транспозосом μ проливает свет на эволюцию рекомбиназы DDE. Природа 491 , 413–417 (2012).
Артикул ОБЪЯВЛЕНИЯ пабмед ПабМед Центральный Google Scholar
Пуджани, А., Рубинштейн, Дж. Л., Флит, Д. Дж. и Брубейкер, М. А. cryoSPARC: алгоритмы для быстрого неконтролируемого определения структуры крио-ЭМ. Нац. Методы 14 , 290–296 (2017).
Артикул КАС пабмед Google Scholar
Маннервик, М. Гены-мишени гомеодоменовых белков. Bioessays 21 , 267–270 (1999).
Артикул КАС пабмед Google Scholar
Хикман, А. Б. и Дайда, Ф. Механизмы транспозиции ДНК. Микробиологический спектр. 3 , 0034–2014 (2015). МДНА3-.
Артикул Google Scholar
Siguier, P., Gourbeyre, E. & Chandler, M. Известные известные, известные неизвестные и неизвестные неизвестные в прокариотической транспозиции. Курс. мнение микробиол. 38 , 171–180 (2017).
Артикул КАС пабмед Google Scholar
Allingham, J.S., Pribil, P.A. & Haniford, D.B. Все три остатка каталитической триады транспозазы Tn 10 DDE функционируют в связывании ионов двухвалентных металлов. Дж. Мол. биол. 289 , 1195–1206 (1999).
Артикул КАС пабмед Google Scholar
Peterson, G. & Reznikoff, W. Мутации активного сайта транспозазы Tn5 указывают на положение донорской основной ДНК в синаптическом комплексе. J. Biol. хим. 278 , 1904–1909 (2003).
Артикул КАС пабмед Google Scholar
Холм, Л. и Лааксо, Л. М. Обновление сервера Дали. Рез. нуклеиновых кислот . 44 , W351–W355 (2016).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Xu, H. E. et al. Кристаллическая структура комплекса парный домен Pax6 человека и ДНК показывает специфическую роль линкерной области и карбоксиконцевого субдомена в связывании ДНК. Гены Дев. 13 , 1263–1275 (1999).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Benler, S. et al. Гены-карго Tn7-подобных транспозонов включают огромное разнообразие защитных систем, мобильных генетических элементов и генов устойчивости к антибиотикам. mBio 12 , e0293821 (2021).
Артикул пабмед Google Scholar
Querques, I., Schmitz, M., Oberli, S., Chanez, C. & Jinek, M. Выбор и ремоделирование целевых участков с помощью систем CRISPR-транспозонов типа V. Природа 599 , 497–502 (2021).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Xiao, R. et al. Структурные основы распознавания ДНК-мишени с помощью CRISPR-Cas12k для РНК-управляемой транспозиции ДНК. Мол. Сотовый 81 , 4457–4466.e4455 (2021).
Артикул КАС пабмед Google Scholar
Park, J. U. et al. Структурная основа для выбора сайта-мишени в системах транспозиции ДНК, управляемых РНК. Наука 373 , 768–774 (2021).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Халпин-Хили, Т.С., Кломпе, С.Е., Штернберг, С.Х. и Фернандес, И.С. Структурная основа нацеливания ДНК с помощью системы CRISPR-Cas, кодируемой транспозонами. Природа 577 , 271–274 (2020).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед Google Scholar
Jia, N., Xie, W., de la Cruz, M.J., Eng, E.T. и Patel, D.J. Структурно-функциональное понимание начального этапа интеграции ДНК с помощью комплекса CRISPR-Cas-Transposon. Сотовый рез. 30 , 182–184 (2020).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Skelding, Z., Queen-Baker, J. & Craig, N.L. Альтернативные взаимодействия между транспозазой Tn7 и белком, связывающим ДНК-мишень Tn7, регулируют иммунитет и транспозицию мишени. EMBO J. 22 , 5904–5917 (2003).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Холодий Г.Ю. и др. Четыре гена, два конца и res-область участвуют в транспозиции Tn5053: парадигма нового семейства транспозонов, несущих либо мероперон, либо интегрон. мол. микробиол. 17 , 1189 (1995).
Артикул КАС пабмед Google Scholar
Николас Э. и др. Семейство репликативных транспозонов Tn3. Микробиолог. Спектр. 3 , 0060–2014 (2014). МДНК3.
Google Scholar
Петровский С. и Станисич В. А. Tn502 и Tn512 являются охотниками за res-сайтами, что свидетельствует о независимой от резольвазы транспозиции в случайные сайты. Дж. Бактериол. 192 , 1865–1874 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Минахина С., Холодий Г., Миндлин С., Юрьева О., Никифоров В. Транспозоны семейства Tn5053 являются охотниками за рез-сайтами, распознающими плазмидные рез-сайты, занятые родственными резольвазами. Мол. микробиол. 33 , 1059 (1999).
Артикул КАС пабмед Google Scholar
Chen, Q. et al. Структурная основа бесшовного вырезания и специфического нацеливания транспозазой piggyBac. Нац. коммун. 11 , 3446 (2020).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Пунджани, А. и Флит, Д. Дж. Трехмерный анализ изменчивости: выявление непрерывной гибкости и дискретной неоднородности крио-ЭМ с одной частицей. Дж. Структура. биол. 213 , 107702 (2021).
Артикул КАС пабмед Google Scholar
Вилас, Дж. Л. и др. MonoRes: автоматическая и точная оценка локального разрешения для карт электронной микроскопии. Структура 26 , 337–344.e334 (2018).
Артикул КАС пабмед Google Scholar
Санчес-Гарсия, Р. и др. DeepEMhancer: решение для глубокого обучения для крио-ЭМ объемной постобработки. Комм. биол. 4 , 874 (2021).
Артикул пабмед ПабМед Центральный Google Scholar
Bienert, S. et al. Репозиторий SWISS-MODEL — новые возможности и функциональность. Рез. нуклеиновых кислот. 45 , Д313–д319 (2017).
Артикул КАС пабмед Google Scholar
Goddard, T.D. et al. UCSF ChimeraX: решение современных задач визуализации и анализа. Науки о белках. 27 , 14–25 (2018).
Артикул КАС пабмед Google Scholar
Эмсли П., Локамп Б., Скотт В. Г. и Коутан К. Особенности и развитие Coot. Acta Кристаллогр. Д. биол. Кристаллогр. 66 , 486–501 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Бэк, М. и др. Точное предсказание белковых структур и взаимодействий с использованием трехдорожечной нейронной сети. Наука 373 , 871–876 (2021).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Адамс, П. Д. и др. PHENIX: комплексная система на основе Python для решения макромолекулярной структуры. Acta Кристаллогр. Д. биол. Кристаллогр. 66 , 213–221 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Kidmose, R. T. et al. Namdinator — автоматическая молекулярно-динамическая гибкая подгонка структурных моделей к крио-ЭМ и кристаллографическим экспериментальным картам. IUCrJ 6 , 526–531 (2019).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Сайто, М. и др. Двойной режим CRISPR-ассоциированного самонаведения транспозонов. Cell 184 , 2441–2453.e2418 (2021).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Загрузить ссылки
Благодарности
Мы благодарим Датский национальный центр крио-ЭМ в CFIM Копенгагенского университета и особенно Тиллманн Пейп за поддержку во время сбора крио-ЭМ данных. Мы также благодарим отдел экспрессии белков CPR за помощь в экспрессии и очистке белков. ФТК является участником Копенгагенской программы докторантуры по биологическим наукам (NNF19SA0035440). Г.М. является частью Центра белковых исследований (CPR) Фонда Ново Нордиск, который финансируется Фондом Ново Нордиск (грант NNF14CC0001). Эта работа также была поддержана грантом NNF0024386, грантом NNF17SA0030214 и грантами Distinguished Investigator (NNF18OC0055061) для GM, который является членом Кластера интегративной структурной биологии (ISBUC) в Копенгагенском университете.
Информация об авторе
Примечания автора
Стефано Стелла
Текущий адрес: Twelve Bio ApS, Ole Maaløes Vej 3, 2200, Копенгаген, Дания
Авторы и филиалы
Группа структурной молекулярной биологии, Фонд Ново Нордиск, Центр белковых исследований, Университет здравоохранения и наук of Copenhagen, 2200, Копенгаген, Дания
Франциско Тенхо-Кастаньо, Николас Софос, Луиза С.
Штуцке, Андерс Фульсанг, Стефано Стелла и Гильермо МонтойяОтделение по очистке и характеристике белков, Исследовательский центр Ново Нордиск, Факультет Копенгагенского университета здоровья и медицинских наук, 2200, Копенгаген, Дания
Blanca López-Méndez
Авторы
- Francisco Tenjo-Castaño
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Николас Софос
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Blanca López-Méndez
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Luisa S. Stutzke
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Anders Fuglsang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Stefano Stella
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Гильермо Монтойя
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
G. M. задумал исследование, F.T.C. и Г.М. поставил биохимические эксперименты. ФТК и С.С. установили протокол экспрессии и очистки. ФТК и AF охарактеризовали свойства связывания белок-ДНК. ФТК и Л.С. создали мутантов и провели эксперименты по транспозиции. ФТК и Б.Л.М. выполнил анализ SEC-MALS, F.T.C. подготовили сетки ЭМ и собрали крио-ЭМ изображения с Н.С. ФТК и Н.С. выполнил крио-ЭМ обработку данных, построил модели и Г.М. приступил к крио-ЭМ карте и анализу структуры с их помощью. Общие результаты обсуждались и оценивались со всеми авторами. Г.М. координировал и руководил проектом и написал рукопись с участием всех авторов.
Автор, ответственный за переписку
Переписка с Гильермо Монтойя.
Декларации этики
Конкурирующие интересы
Г.М. и S.S. являются соучредителями и членами Twelve Bio BoD. Остальные авторы заявляют об отсутствии конкурирующих интересов.
Рецензирование
Информация о рецензировании
Nature Communications благодарит анонимных рецензентов за их вклад в рецензирование этой работы. Доступны отчеты рецензентов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Файл рецензирования
Описание дополнительных дополнительных файлов
Дополнительный фильм 1
.0005
Исходные данные
Исходные данные
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате. , при условии, что вы укажете первоначальных авторов и источник, предоставите ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
VK 45.02 A — Немецкий тяжёлый танк VIII уровня
Blitz Ангар
База танков и советы по World of Tanks Blitz
Если вам нравится проект, поддержите его на Патреон или взгляните на наш Магазин товаров
Стоимость
- 2 435 000
- 90 000
Предшественник
- VIIТигр (P)
Преемник
- IXВК 45. 02 Б
Конкурирует с
- VIIIТигр II
- VIIIВК 100.01 (П)
Конфигурация
Провизия
Оборудование
Модули
Навыки экипажа
Расходные материалы
- 90-е годы
- 90-е годы
- 90-е годы
- 75 с
15 с
- 90-е годы
- 75 с
15 с
- 90-е годы
20-е годы
Бустеры
Параметры
Сравните параметры танка с другими из:
1750
HP
30 / 45
282,5 м
Дальность обзора
2 / 45
Огневая мощь
2270
ДПМ
15 / 45
215
Penetration
37 / 45
310
Damage
26 / 45
150
Devices/crew damage
22 / 45
—
Burst damage
950
Velocity
28 / 45
9,30%
Потеря проникновения на расстоянии
14 /45
—
Радиус взрыва
1904
DPM
15 /44
265
ПЕРЕДЕЛЕНИЕ
. 0006 19 /44260
повреждение
26 /44
150
Устройства / урон экипажа
21 /44
—
. %
Потеря проникновения на расстоянии
12 /28
—
Радиус взрыва
3076
DPM
9 /44
60
. Проникновение
18 /44
42059
.0005
повреждение
23 /44
150
Устройства / повреждение экипажа
21 /44
—
Повреждение.
1,91
Радиус взрыва
22 /44
Проникновение
Повреждение
Устройства / ущерб экипажу
Потерян.0005
Explosion radius
105.0
Caliber
24 / 45
7.32
Rate of fire
14 / 45
8.19
Reload time
10 / 36
—
Magazine reload time
Время заряжания следующего снаряда
—×
Размер магазина
—
Время до готовности следующего снаряда
Обращение с оружием
2,08
Базовое время сведения
8 / 45
0. 335
Dispersion
15 / 45
Dispersion factors
0.160
When moving
3 / 45
0.160
When rotating
4 / 45
0.100
При вращении башни
2 /45
3.000
после выстрела
1 /45
Углы оружия
6 °
Депрессия
31 /45
17 °
Элевация
31 /4517 °
660005
26/45
180 ° / 180 °
Арма (слева / справа)
1 /45
29,16
Трорачная скорость
15 /45
Мобильность
565
9699959
9
9
9
9
9
9
99965
9
9
5
699699
9599999999999999999999999999599
26 / 45
38
Вперед
20 / 45
16
Назад
10 / 45
Эффективная мощность/масса
14,76
На пересеченной местности
7 / 45
12. 49
On medium terrain
4 / 45
6.49
On soft terrain
34 / 45
Traverse speed
44.82
On hard terrain
1 / 45
37,93
на средней местности
1 /45
19,72
на мягкой местности
13 /45
6,66 м × 3,12 м..23%
, в то время как все еще
34 /45
5,62%
при перемещении
33 /45
3,51%
при стрельбе, пока
35 /45
2,76%
6.
35 / 45
Дистанция обнаружения противником с дальностью обзора 271 м
250,6 м
На месте
258,6 м
На ходу
263,2 м
При стрельбе на месте
264,9 м
При стрельбе в движении
Основная броня
240–300
180–240
120–180
60–120
0–60
Разнесенная броня
288–360
216–288
144–216
72–144
0–72
Голова
Сбоку
Сзади
Инициализация…
Пик-а-бу Средний
- Рассеивание хороший
- Дисперсия в движении хороший
- Соотношение мощность/вес средний
- Скорость назад хороший
- Прицельная дуга хороший
Средний
- Рассеивание хороший
- Дисперсия в движении хороший
- Соотношение мощность/вес средний
- Скорость назад хороший
- Прицельная дуга хороший
Вы прячетесь за укрытием, полностью перезаряжаете свое оружие, предварительно прицеливаетесь и ускользаете
за секунду сделать один поражающий выстрел. Прежде чем враг поймет, что вы
снова спрятался. Достойная подвижность и стабилизация прицеливания и вы
конечно может осуществить это один.
Снайперская стрельба на дальние дистанции Средний
- Рассеивание средний
- Скорость снаряда хороший
- Повреждать хороший
- Проникновение хороший
Средний
- Рассеивание средний
- Скорость снаряда хороший
- Повреждать средний
- Проникновение средний
Прятаться в кустах на холме и стрелять по врагам с другой стороны
карты. Если вы не возражаете против всей ненависти, которая приходит с этим, тогда вы
просто нужно хорошее рассеивание и пробитие, чтобы каждый выстрел был засчитан.
Линия фронта Не рекомендуется
- Передняя броня плохой
Не рекомендуется
- Передняя броня плохой
Забудьте о скалах, холмах, зданиях и прочем укрытии. Сделай себя прикрытие для ваших товарищей по команде. Конечно, это работает, только если у вас очень хорошую броню и уметь ею пользоваться или… ну если ты уже мертв.
Круг смерти Не рекомендуется
- Прицельная дуга хороший
- Скорость перемещения плохой
- Соотношение мощность/вес средний
Не рекомендуется
- Прицельная дуга хороший
- Скорость перемещения плохой
- Соотношение мощность/вес средний
Двигайтесь за медленными истребителями танков и тяжелыми танками и продолжайте кружить
вокруг них, чтобы избежать удара, делая легкие выстрелы в спину
врага. Требуется хорошая скорость перемещения и ускорение.
Ударь и беги Не рекомендуется
- Диапазон просмотра хороший
- Скорость вперед плохой
- Соотношение мощность/вес плохой
Не рекомендуется
- Диапазон просмотра средний
- Скорость вперед плохой
- Соотношение мощность/вес плохой
Спрячьтесь в кустах, незаметно заметьте врага. Сделайте снимок и
начать бежать. Быстрый. Повторяйте, пока не останется врагов. Овладейте этой техникой
и на вашем пути будет много мастерства.
драка 1 на 1 Не рекомендуется
- ДПМ плохой
Не рекомендуется
- ДПМ плохой
Кто победит в драке 1 на 1, зависит от слишком многих переменных. Броня, оружие, подвижность, размеры танков и многое другое. Одно точно: без добра DPM: лучше оставаться в группе или держаться на расстоянии.
Историческая справка
Разработка этой машины началась в апреле 1942 года. Фирма «Крупп» получила заказ на постройку башни. Однако прототип так и не был изготовлен. Башни устанавливались на первые Tiger II.
Reddit — Погрузитесь во что угодно
О чем все это?
В последнее время я заметил несколько случаев, когда утверждалось, что VK-00 является самым быстрым квантовым приводом (QD) размера 1 без особого объяснения, почему. Это включает в себя обсуждения здесь, на Reddit, а также видео создателей контента на Youtube.
Я хотел бы воспользоваться этой возможностью, чтобы показать, почему это утверждение в целом чрезмерно упрощено, с одной стороны, и по большей части ошибочно, с другой.
источник: https://starcitizen.tools/VK-00
[ TLDR]
Параметр «скорость» квантового привода часто неправильно интерпретируется. Правильное значение — максимальная скорость, а не фактическая скорость, на которую также сильно влияют значения ускорения. Вместо этого используйте стелс-драйв, если хотите как можно быстрее преодолевать короткие и средние расстояния.
Как получилось?
У меня есть сильное подозрение относительно того, как возникает это предположение. Поскольку игра не предоставляет никакой статистики по внутриигровому QD или любому другому компоненту в этом отношении, люди по понятным причинам обращаются к партийным веб-сайтам 3 rd , таким как Erkul, с намерением принять более взвешенное решение о том, какие компоненты получить. К сожалению, с QD это не так просто, как прямое сравнение параметров. Но это именно то, что, скорее всего, произойдет. Сортировка дисков S1 по «скорости» и ВК-00 оказывается на первом месте…
Модель под капотом
Обозначение этого параметра немного неудачное. Максимальная скорость (vmax) была бы более подходящей. Это максимальная скорость, которую может развить привод. Однако это значение не достигается мгновенно. Сначала привод проходит фазу ускорения. Как видно ниже, скорость, с которой привод разгоняется, не является постоянной, а представляет собой линейную функцию с начальным (a1) и конечным (a2) значениями. Эркул называет их ускорением стадии 1 и стадии 2. Следовательно, это приводит к квадратичному приращению скорости до тех пор, пока привод не достигнет своего значения vmax. Прежде чем достичь места назначения, привод проходит фазу торможения, которая следует тем же правилам.
Вот график, иллюстрирующий это: (переход CRU->HUR)
Рис. 1: Кривые ускорения, скорости и расстояния при переходе CRU->HUR
Итак, что все это значит?
Это означает, что время, необходимое КТ для прохождения определенного расстояния, определяется тремя параметрами, а не только одним. Это также означает, что, поскольку функция зависимости расстояния от времени прохождения не линейная и не квадратичная, а кубическая (!) в принципе невозможно достоверно судить о производительности КТ, глядя на параметры на глаз или только сортируя. Время в пути должно быть рассчитано для каждого интересующего расстояния, поскольку чаще всего водитель может быть быстрее на более короткие расстояния по сравнению с конкурентом, но медленнее на более длинные расстояния или наоборот.
VK-00 против конкурентов
Итак, давайте посмотрим, как VK-00 работает против популярного привода Atlas и невидимого привода Spectre на примере прыжка на 1 км.
| Drive | a1 [km/s 2] | a2 [km/s 2] | vmax [km/s] |
|---|---|---|---|
| VK-00 | 625 | 3’724 | 283’046 |
| Spectre | 1’035 | 3’426 | 201’112 |
| ATLAS | 735 | 2’433 | 151’951 |
Fig. 2: VK-00. VS ATLA и Specster, 1 MK. ускоряется быстрее, чем VK-00 в любой момент времени, и, поскольку это всего лишь короткий скачок, ни один из приводов даже близко не приближается к своему vmax. Так что VK-00 не может играть на своей кажущейся силе, у него высокий vmax. Забавно, но привод с самой высокой пиковой скоростью в этом сценарии — это Spectre. При таких обстоятельствах неудивительно, что он совершает прыжок примерно на 10 секунд быстрее, чем VK-00.
Даже Atlas, с намного более низкими a2 и vmax, по-прежнему совершает прыжок примерно на 2 секунды раньше, чем VK-00.
Таблицы поиска времени в пути
Итак, как это работает для различных расстояний? Ниже приведены три диаграммы лучших дисков S1 (мой личный выбор). Они отображают расстояние в зависимости от времени в пути, разделенные на графики в масштабе ближнего, среднего и дальнего действия для лучшей читаемости.
Рис. 3: Время в пути в зависимости от расстояния, подборка лучших дисков S1
Глядя на эти графики, основное утверждение можно определенно опровергнуть по большей части.
VK-00 на самом деле является самым медленным приводом в группе до 3 Мкм, после чего он начинает опережать второй самый медленный в линейке Atlas.
На последнем графике хорошо видно, что высокое значение vmax, которое часто считают отличительной чертой VK-00, начинает значительно сокращать время в пути только при более длинных прыжках. Однако высокий расход топлива сразу компенсирует это для большинства небольших кораблей с их ограниченными топливными баками. Они иссякают до того, как преимущество действительно начинает проявляться.
Тем не менее, есть несколько кораблей с большими топливными баками, таких как Eclipse или Defender, которые могут довести его до точки, где VK-00 действительно станет самым быстрым S1 QD. Точка разрыва, где он превосходит Spectre, находится прибл. 37 кмкм, после сжигания 797 л Qt топлива:
расстояние зависит от трех параметров. Не только один.
Нет однозначного ответа на упрощенный вопрос о том, какая QD самая быстрая. Это зависит от вашего топливного бака и, прежде всего, от вашего варианта использования и от того, какое расстояние вы хотите преодолеть. Правильный вопрос должен звучать так: какой QD быстрее всего преодолевает XY км?
Поскольку кривая времени нелинейна, лучше всего рассчитать время в пути для интересующего вас расстояния (например, с помощью встроенного калькулятора на Erkul) и непосредственно сравнить результаты. Или используйте диаграммы, чтобы найти это, как те, которые я представил здесь.
Я надеюсь, что это было информативно и что один или другой могли узнать кое-что по пути.
Дополнительную информацию по этой теме можно найти здесь. В документе объясняется весь процесс того, как я вывел модель и математические формулы, которые использовались для создания этих диаграмм, около полутора лет назад.
Cheers, Erec
Заявление об ограничении ответственности
Термин «время в пути» относится к простому времени в пути, без учета времени спулинга или калибровки. Текущим патчем на момент написания была Alpha 3.17.2.
Hootsuite Интеграция веб-приложения с ВКонтакте • Сделано Pioneers
Hootsuite — имя, известное каждому, кто работает в сфере маркетинга в социальных сетях. Более двухсот гордых сотрудников Hootsuite внесли свой вклад в создание самого передового инструмента маркетинга в социальных сетях в Интернете. Будучи тесно интегрированным с ограниченным числом социальных сервисов, Hootsuite также предлагает так называемый «Каталог приложений». Каталог приложений призван привлечь более широкий выбор интеграций с социальными службами, созданных сторонними разработчиками.
ВКонтакте — самая сложная и широко используемая социальная сеть в Европе. По мере того, как инициативы Hootsuite в области онлайн-маркетинга и социальных сетей быстро набирали обороты в регионе, поддержка Hootsuite была переполнена запросами от крупных и средних предприятий, которым требовалось решение с поддержкой ВКонтакте. Вот почему команда Hootsuite решила стать партнером Pioneers Software Inc.
Нам не стыдно признаться, что проект был непростым. Никто никогда не видел веб-клиент ВКонтакте, и требуемый уровень интеграции был намного глубже, чем предполагалось для веб-сайтов. Поэтому требовалось более продвинутое решение, и в результате тесного сотрудничества с командой ВКонтакте нам был предоставлен более глубокий доступ к API так, как никогда раньше. Приложение, которое мы создали вместе с Hootsuite и ВКонтакте, можно считать первым полнофункциональным веб-клиентом ВКонтакте.
Так как приложение обслуживает десятки тысяч пользователей, размещение сервера-посредника между Hootsuite и VK было бы неэффективным. Вместо этого наш подход в этом случае заключался в том, чтобы обеспечить безопасное выполнение запросов API на стороне клиента — это сделало приложение намного более эффективным. Среди технологий интеграции, которые мы использовали, были Ruby on Rails, KnockoutJS и OAuth.
Этот абстрактный поток описывает взаимодействие между пользователями Hootsuite, сервером приложений и VK API.
+--------+ +-------------+ +--------+
| |--(A)- Запрос авторизации -->| | | |
| Клиент | | Сервер приложений | | API ВКонтакте |
| | | |--(B)- Запрос авторизации -->| |
| | | | | |
| | | |<-(C)- Диалог авторизации --| |
| | | | | |
| |<-(D)- Диалог авторизации --| | | |
| | | | | |
| | | |
| |--(E)--------------------- Диалог авторизации -------------------- ------>| |
| | | |
| | | | | |
| | | |<-(F)----- Маркер доступа --------| |
| | | | | |
| |<-(G)----- Маркер доступа --------| | | |
| | +------------+ | |
| | | |
| |--(H)------------- Маркер доступа --------------- ---------------->| |
| | | |
| |<-(I)-------------------------------------- Защищенный ресурс ------------------ -----------| |
+--------+ +--------+
В конце цикла разработки мы провели всестороннее тестирование обеспечения качества (QA) и стресс-тестирование приложения, чтобы убедиться, что оно может работать ожидаемые нагрузки.