
Help — Подача объявлений
Как подать объявление?
Чтобы подать объявление на сайте:
- авторизуйтесь — нажмите кнопку Войти в правом верхнем углу и выберите удобный способ;
- после авторизации подайте объявление, нажав на кнопку Разместить объявление (справа сверху) и заполнив все поля Подробнее
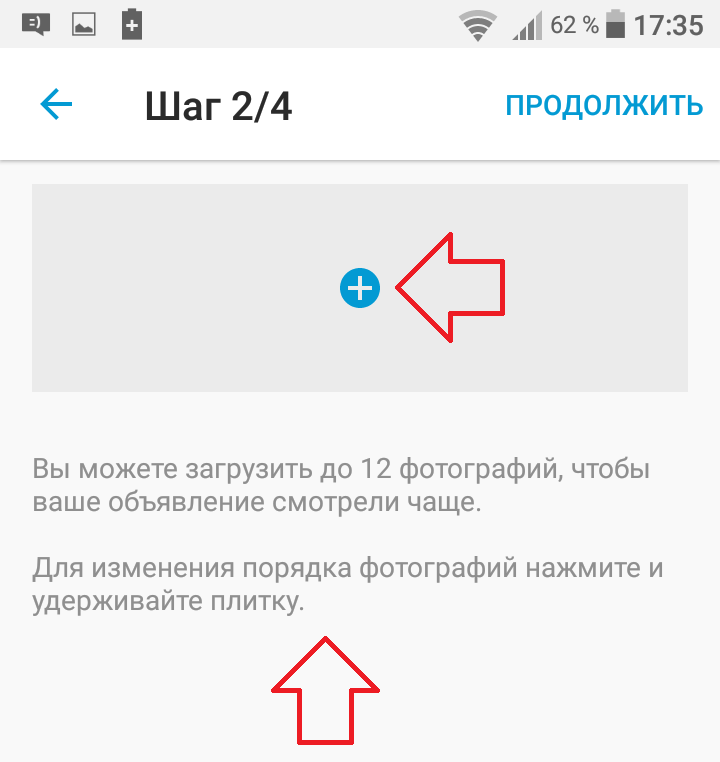
Если вы используете мобильное приложение Юла:
- нажмите на «+» далее авторизуйтесь, если вы еще не вошли в свой профиль
- если вы уже авторизованы, размещайте объявление, заполнив все поля Подробнее
Как массово загружать объявления на Юлу?
В настоящее время такая возможность на Юле доступна лишь в рамках размещения через Тариф. Данная услуга позволяет совершать различные массовые операции с действиями, в том числе, и подачу. Подробнее о тарифах
Размещение объявлений на Юле — платное или бесплатное?
На Юле есть лимиты бесплатных объявлений – это максимальное количество публикаций, которое вы можете размещать бесплатно в течение 30 дней.
Для категорий Вакансии, Для бизнеса, Спецтехника (подкатегории: Автобусы и грузовики и Спецтехника) и Коммерческая недвижимость бесплатных размещений нет. Полный список лимитов по категориям
Размещать объявления сверх бесплатного лимита можно на платной основе. Подробнее о пакетах размещений
Какие правила размещения объявлений есть на Юле?
Чтобы ваше объявление успешно прошло модерацию, рекомендуем следовать правилам нашего сервиса. Вы можете ознакомиться с ними здесь:
- общие требования к размещению объявлений
- требования к описанию товара и услуги
- требования к фотографиям
Все правила сервиса
Какие товары и услуги нельзя размещать в объявлениях на Юле?
На нашем сервисе, есть ряд товаров и услуг, запрещенных к продаже. Чтобы ваше объявление успешно прошло модерацию, изучите запрещенные товары и услуги здесь.
Можно ли добавлять в объявления видео или ссылки на них?
В настоящий момент на Юле нет возможности добавлять видео.
Указывать в объявлении ссылки на сторонние ресурсы запрещено правилами нашего сервиса. Требования к описанию товара или услуги
Можно ли подавать одинаковые объявления по разным городам или адресам?
Размещать одинаковые объявления с разным местоположением можно только в категории «Вакансии».
Во всех остальных категориях для одного товара или услуги должно быть размещено только одно объявление.
Общие требования к размещению объявлений
2. Запрещено размещение одинаковых или схожих объявлений, независимо от указанного местоположения или выбранной категории, в том числе через разные аккаунты.
В какой категории нужно размещать объявления об услугах?
Если вы хотите опубликовать объявление как исполнитель, то вам необходима категория Услуги исполнителя/Предложить свои услуги.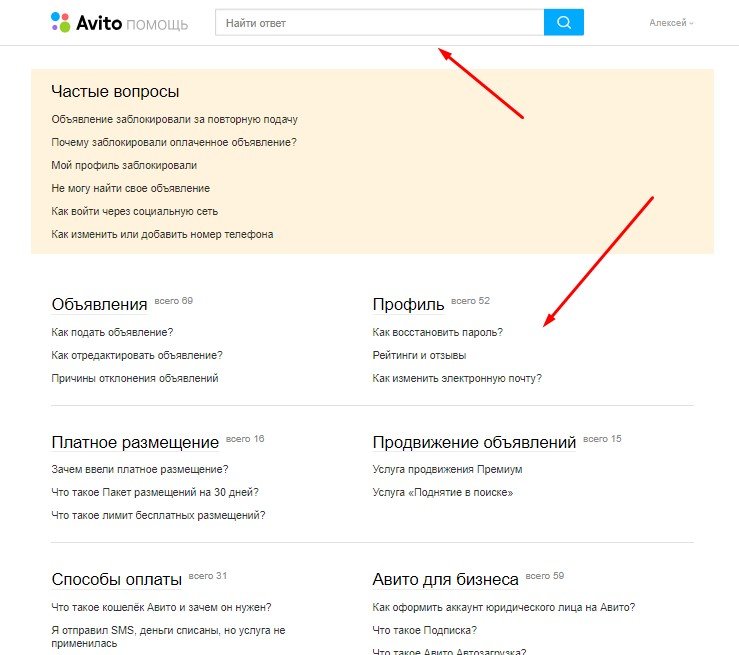 Подробнее
Подробнее
Если вы ищете специалиста, который выполнит услугу для вас, выбирайте категорию Заявки на услугу / Заказать услугу. Подробнее
Какой оптимальный размер фотографий, которые можно разместить в объявлениях?
Допустимый размер фотографий для публикации объявления не более 25мб. Они должны быть в стандартном формате JPG или PNG. Фотографии которое весят больше, к сожалению, не могут быть загружены технически.
Если у вас фотографии бОльшего размера или имеют другой формат
, воспользуйтесь каким-либо редактором для сжатия/форматирования изображений.Как сделать так, чтобы объявления показывались по всей России?
Объявление доступно любому пользователю из любого города, однако, геотег (город) в фильтрах поиска должен совпадать с геотегом объявления. Увидит ли пользователь из другого города ваше объявление зависит только от настроек радиуса его поиска.
Чтобы шансы на быструю продажу увеличились, вы можете:
- включить доставку в объявлении, и отправлять свои товары через нашу доставку Боксберри. Подробнее о доставке
- подключить услугу продвижения по всей России. Тогда ваше объявление будет периодически показываться в ленте в других городах. Подробнее об услугах продвижения
Как заново опубликовать объявление из раздела Продано?
Если сделка сорвалась, и вы снова хотите продавать товар, ранее отмеченный как Продано (или у вас появился новый товар, похожий на уже проданный), вам нужно создать новое объявление с новыми фотографиями. Дело в том, что система запоминает все загруженные в нее тексты и фото, даже проданные и удаленные, поэтому дубликат объявления может быть заблокирован. Подробнее
Как скопировать ссылку на свое объявление?
Чтобы скопировать ссылку на объявление в приложении Юла, внизу под описанием объявления нажмите на кнопку с двумя прямоугольниками на голубом фоне.
Ссылка будет скопирована в буфер памяти. Перейдите в диалог, куда вы хотите отправить ссылку, нажмите долгим тапом на поле для ввода текста и выберите опцию Вставить.
Возможные трудности при публикации объявления
При попытке опубликовать объявление появляется ошибка или приложение вылетает
Если вы используете мобильное приложение Юла, обновите его до последней версии:
- Android
- iOs
Также попробуйте стереть данные приложения (Настройки телефона -> Приложения -> ЮЛА -> Стереть данные) и очистить кэш .
Если вы используете сайт Юлы, сделайте очистку cache и cookies браузера.
При попытке разместить объявление появляется ошибка «Превышен времени запроса при размещении объявления»
Ошибка превышения времени запроса при размещении объявления чаще всего возникает из-за плохого качества связи в данный момент. Попробуйте сменить способ подключения к интернет с wi-fi на сотовую связь или наоборот. Повторите попытку размещения объявления через некоторое время.
Попробуйте сменить способ подключения к интернет с wi-fi на сотовую связь или наоборот. Повторите попытку размещения объявления через некоторое время.
Не загружаются фотографии в объявление
Если вы используете сайт Юлы, сделайте очистку cache и cookies браузера.
Если проблема возникает в мобильном приложении, то попробуйте стереть данные приложения (Настройки телефона -> Приложения -> ЮЛА -> Стереть данные) и очистить кэш.
При попытке разместить фото появляется ошибка «Неподдерживаемый тип файла»
Если вы используете сайт Юлы, сделайте очистку cache и cookies браузера или попробуйте использовать другой браузер.
Если проблема возникает в мобильном приложении, то попробуйте стереть данные приложения (Настройки телефона -> Приложения -> ЮЛА -> Стереть данные) и очистить кэш.
При публикации объявления фотографии переворачиваются
Чтобы изображения не переворачивались, вам нужно уменьшить фото до допустимого размера (не больше 25мб) и обрезать их до квадрата.
Не могу выбрать в объявлении нужный адрес
Мы используем карты Google. Поэтому нужный адрес или объект там есть и относится к территории Российской Федерации, а в приложении Юла отсутствует, пришлите нам ссылку на него в Google Maps.
Если объект отсутствует на картах Google, или же по какой-то причине Google относит его к территории другого государства (не РФ), рекомендуем обратиться в службу поддержки этого сервиса через меню слева вверху на их сайте.
В объявлении указывается другое местоположение
Если вы пользуетесь мобильном приложением Юла, то в разделе Мой профиль — Настройки есть пункт Личные данные. Если вы в нем укажите адрес, то при публикации новых объявлениях в мобильном приложении этот адрес вам и будет предлагаться.
На сайте же это работает немного по-другому. Нужно указать адрес не в настройках профиля, а разделе Местоположение, которое находится в самом верху сайта. После указания адреса в указанном разделе, адрес будет указывать при размещении новых объявлений.
Адрес не закрепляется и автоматически меняется на другой
Если вас не устраивает адрес, подставленный автоматически, или же вас определяют в месте, где адрес невозможно установить автоматически, необходимо указать нужный адрес вручную.
При ручном вводе адреса напишите его без лишних символов (например, «Орёл Ленина 5» или «орел ленина 5») и немного подождите – должен появиться выпадающий список, в котором вы можете выбрать нужную строку.
В объявлении мало просмотров
Количество просмотров, частота продажи напрямую зависят от объявления: описание, фото, категория, гео, востребованность, даже сезонность и т.д.. Объявления видят все пользователи, но повлиять на то, захочет ли пользователь просмотреть объявление и купить товар, мы не можем.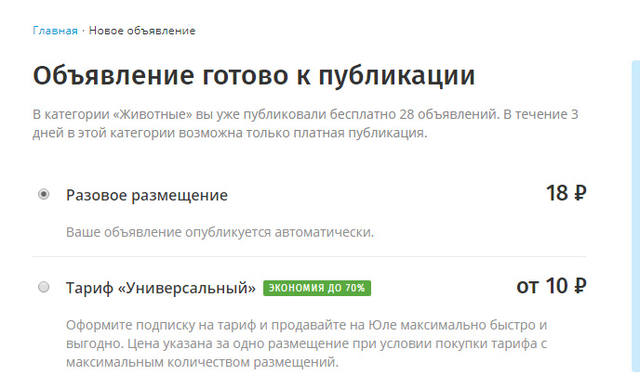
Покупатели могут проявить интерес к вашему объявлению, если оно будет актуально и привлекательно. Поэтому если вам кажется, что просмотров у объявления мало, попробуйте изменить название, цену или сделать более привлекательные фотографии (особенно, первую – ведь именно ее видят пользователи в ленте).
В объявлении нет кнопок Позвонить и Написать
Кнопки Позвонить и Написать действительно отсутствуют в объявлениях у самого продавца.
Но они всегда доступны другим пользователям. Чтобы убедиться в этом, вы можете проверить свои объявления из другого аккаунта. Подробнее
Вопрос по редактированию объявления
Вопрос по снятию с публикации и восстановлению объявления
Support
We will help solve the problem
«Юла» запустила возможность продвигать товары с помощью видео
Продолжение сюжета от
Новости СМИ2
Новости
Новости
Анастасия Марьина
Руководитель новостного отдела RB. ru
ru
Анастасия Марьина
Сервис объявлений «Юла» (принадлежит Mail.ru Group) добавил возможность загружать на ресурс видео про товары и услуги, которые продают пользователи. Об этом говорится в пресс-релизе сервиса.
Анастасия Марьина
Продавцы и мастера смогут бесплатно продвигать свои объявления и профили с помощью коротких видеороликов в «историях», работающих на базе технологий видеоплатформы соцсети «Одноклассники».
Весь контент будет проходить модерацию, а видео можно будет снять на камеру или добавить из уже имеющихся медиафайлов.
Изображения: пресс-служба «Юлы»
Ранее в «историях» на «Юле» можно было размещать только фотографии, прикрепив к ним ссылку на объявление или профиль пользователя.
В отличие от социальных сетей на Юле «истории» видны в течение недели и показываются не только подписчикам продавца, но и другим пользователям сервиса. В начале списка «историй» пользователь видит контент от продавцов, на которых он подписан, а затем от тех, кто находится неподалеку.
В начале списка «историй» пользователь видит контент от продавцов, на которых он подписан, а затем от тех, кто находится неподалеку.
- Реклама
- Полезные сервисы
- Бизнес
- VK (корпорация)
- Видео
Нашли опечатку? Выделите текст и нажмите Ctrl + Enter
Материалы по теме
- 1 5 неочевидных замен Youtube для бизнеса
- 2 Работа над инновационным продуктом за пять шагов
- 3 Альтернативные инвестиции: что это и как они работают
- 4 Автомобильный бизнес: как продавать эффективно?
- 5 На каких tech-креаторов подписаться в российском TikTok?
ВОЗМОЖНОСТИ
15 сентября 2022
SechenovTech
15 сентября 2022
Food Up
15 сентября 2022
Акселератор РЖД
Все ВОЗМОЖНОСТИ
Новости
«Яндекс» представил цифровую карту с кешбэком и подключенным Mir Pay
Колонки
Как переводить деньги в Европу из России в 2022 году?
Новости
Эксперт «Купибилет» объяснила, как россияне смогут путешествовать на Кипр по национальной визе
Колонки
9 перспективных бизнес-идей после ухода иностранных компаний
Новости
Business Insider: Маск отложил покупку Twitter из-за речи Путина к 9 мая
Как заработать на Юле: получаем стартовый капитал
Юла – молодой сервис бесплатных объявлений. Отличия от Авито заключаются в модерации – на Юле она более лояльная. Кроме того, допускается автоматическое размещение объявлений. Заработать на первоначальный капитал при помощи сервера youla.ru можно, работая по системе дропшиппинга или партнерским программам. Подробно расскажем, как это сделать.
Отличия от Авито заключаются в модерации – на Юле она более лояльная. Кроме того, допускается автоматическое размещение объявлений. Заработать на первоначальный капитал при помощи сервера youla.ru можно, работая по системе дропшиппинга или партнерским программам. Подробно расскажем, как это сделать.
Заработать на Юле на первоначальный капитал можно двумя распространенными способами: торговлей по системе дропшиппинга и при помощи партнерских программ.
Дропшиппинг
Продажа товаров по системе дропшиппинга предполагает, что вы занимаетесь только поиском покупателя, а все остальные действия (такие как упаковка и отправка заказа и работа с покупателем) осуществляются дропшиппинговой платформой (поставщиком). Прямая поставка, или Дропшип (от англ. dropshipping), — это метод управления цепью поставок, при которой посредник осуществляет розничную торговлю без промежуточного хранения товаров на своём складе.
Интересная информация! Заработать деньги на первоначальный капитал, а также обеспечить себе пассивный доход можно, размещая видео ролики на YouТube.
Самыми популярными площадками дропшиппинга, работа на которых интуитивно понятна и проста, являются:
- qnits.ru;
- cdrop.ru;
- insales.ru;
- rusdropshipping.ru.
Преимущества площадок для дропшиппинга:
- возможность самостоятельной установки цены выше определенного предела;
- широкий ассортимент;
- отсутствие необходимости организации склада и доставки;
- простое отслеживание заказов, легкий вывод средств.
Этапы работы
Рассмотрим подробно, как заработать на доске бесплатных объявлений «Юла» при помощи дропшиппинга на примере площадки qnits.ru.
Вначале надо будет зарегистрироваться на площадке дропшиппинга, а затем зайти на «Юлу».
Регистрация на сайте дропшиппинга поставщика и выбор изделий для продажи:
Регистрация на Юле и размещение товара:
Продажа товара
После звонка или письма покупателя у него нужно узнать его номер телефона, имя и фамилию. Эти сведения передаются поставщику путем оформления заказа.
Рис. 10. Создание заказа на дропшиппинг-площадке
В открывшейся форме необходимо заполнить количество и цену изделия. Стоимость может быть ниже указанной изначально, но только за счет вашего бонуса. Она может быть и выше. Превышение пойдет вам в бонус. Поставщик заберет себе только указанную им цену, без учета бонуса.
Нижние три строчки заполняйте данными клиента.
Рис. 11. Заполнение бланка заказа
После поступления денег от клиента бонус будет зачислен на ваш счет.
Партнерские программы
По сути, это тот же метод, что и дропшиппинг, но выбрав товар для продажи, вы получаете на него ссылку, которую должны предоставить покупателю, и уже он, перейдя по ссылке, оформляет заказ. Если вы хотите держать все под строгим контролем и оформлять заказы самостоятельно, то необходимо обеспечить поступление заказов с различных IP-адресов. Для этого можно воспользоваться сторонними сервисами, на которых сформировать задание по заполнению заказов, предоставленными вами данными.
Наиболее популярные партнерские программы:
- ad1.ru;
- funcpa.ru;
- kma.biz;
- admitad.com.
Интересная информация! Продавать товары можно и на Одноклассниках. Сайт помогает обеспечить как активный, так и пассивный доход.
Чтобы заработок на Юле приносил максимальные результаты, необходимо размещать большое количество объявлений по всей России. Не стоит зацикливаться на больших городах. В регионах продажи могут быть даже эффективнее. Для автоматизации размещения объявлений можно воспользоваться сервисом y-bot.ru.
Количество объявлений на сайте Юла ограничено.
Категория товара | Количество объявлений (бесплатных) |
|---|---|
Детские товары | 90 |
Детская одежда | 92 |
Хобби и развлечения | 90 |
Запчасти и товары для авто | 10 |
Женская одежда | 90 |
Красота и здоровье | 10 |
Спорт и отдых | 90 |
Мужская одежда | 90 |
Для дома и дачи | 9 |
Компьютерная техника | 90 |
Бытовая техника | 90 |
Телефоны и планшеты | 5 |
Источник: помощь интернет портала mail. ru
ru
Старайтесь размещать максимальное количество бесплатных объявлений. Для поднятия дохода можно воспользоваться пакетами размещений. Сервер предлагает:
- персональные скидки до 70%;
- автоматическое продление размещения объявлений;
- автоматическое поднятие объявлений, что повышает их рейтинг.
Однако на начальном этапе не стоит делать вложения. Бесплатное объявление будет показываться на сайте в течение 30 дней. За это время уже поступит первый доход, и тогда полученные средства можно вложить в автоматизацию.
Не забывайте продвигать выбранные товары в социальных сетях. Это поможет привлечь дополнительное количество клиентов. Вы можете размещать рекламные посты с описанием продукции.
Как создать объявление на eBay. Инструкция, как разместить товар
Откройте Карточку создания объявления и следуйте этой инструкции. Кнопку Создать объявление (Create listing) можно найти в Центре продавцов (Seller Hub) на вкладке Объявления (Listings), в разделе Мой eBay или в правом верхнем углу любой страницы на сайте export. ebay.com.
ebay.com.
Примечание: если вы хотите сделать объявление максимально эффективным, прочитайте рекомендации по оформлению.
Инструкция по созданию объявлений доступна в видео- и текстовом форматах.
Пошаговая инструкция: текстовая версия
Откройте Карточку создания объявления
Откройте Карточку создания объявления в соседнем с инструкцией окне, чтобы было проще следовать подсказкам. Обратите внимание: она доступна только на английском языке, и заполнять ее также нужно по-английски.
Введите название или выберите категорию товара
На главной странице Карточки вы увидите заголовок Расскажите о своем товаре (Tell us what you’re selling). Под ним расположено поле ввода с подсказкой “Укажите UPC, ISBN, ePID, код детали или название продукта”.
У вас есть несколько вариантов:
1. Можно указать название, категорию или идентификатор вашего товара в поле ввода (на английском языке). eBay предложит вам несколько подходящих категорий. Вы можете выбрать одну из них или же кликнуть на первую в списке опцию Создать объявление с этим заголовком (Start with this title). Вы будете автоматически перенаправлены к форме для заполнения информации о товаре.
Вы можете выбрать одну из них или же кликнуть на первую в списке опцию Создать объявление с этим заголовком (Start with this title). Вы будете автоматически перенаправлены к форме для заполнения информации о товаре.
2. Также вы можете выбрать категорию из списка (Browse categories). Нажмите на соответствующую кнопку под полем ввода, а затем укажите подходящую для вашего товара категорию. При выборе открывается список подкатегорий. Как только выбрана окончательная подкатегория, вы автоматически перейдете к форме для заполнения информации о товаре.
Примечание: Если у вашего товара есть цифровой идентификатор, это заметно ускорит создание объявления (категория, параметры и характеристики будут загружены автоматически). Больше о цифровых идентификаторах можно прочитать здесь.
Примечание: Если вы продаете однотипные товары со схожими параметрами, объедините их в одном объявлении с вариациями (Listing with variations). Эта возможность доступна не во всех категориях. Подробнее об Объявлениях с вариациями читайте здесь.
Подробнее об Объявлениях с вариациями читайте здесь.
Заполните Информацию о товаре (Listing details)
Расскажите о своем товаре, предоставив максимум информации. Обратите внимание на то, что некоторые дополнительные опции оформления являются платными (например, Подзаголовок или выделение полужирным шрифтом).
1. В поле Заголовок (Title) введите информативный заголовок длиной до 80 символов. Попробуйте представить, по каким словам или фразам покупатели будут искать ваш товар. Заголовок не обязательно должен выглядеть как связное предложение. Поэтому вам не нужно использовать в заголовке предлоги, союзы и знаки препинания, чтобы не тратить впустую лимит символов.
2. Если нужно, скорректируйте категорию товара с помощью опции Изменить категорию (Change category). Помните, что от правильно выбранной категории зависит продажа вашего товара. При необходимости вы можете добавить вторую категорию (Add a second category), но за размещение товара в двух категориях могут применяться сборы.
3. В поле Состояние (Condition) выберите подходящий пункт из выпадающего меню: Новое с бирками (New with tags), Новое без бирок (New without tags), Новое с дефектами (New with defects), Бывшее в употреблении (Pre-owned).
4. В разделе Фотографии (Photos) добавьте не более 12 фото вашего товара. Их можно загрузить с устройства или импортировать из веб-ресурса (Import from web). Старайтесь размещать максимальное количество фотографий: чем их больше, тем выше шанс найти покупателя. Прочитайте, как правильно сфотографировать товар для eBay.
5. Содержание раздела Характеристики товара (Item specifics) зависит от категории товара. Это может быть бренд, размер, подвид, цвет, стиль или иная полезная для покупателей информация.
Характеристики разделены на три группы: Обязательные (Required), Рекомендуемые (Recommended) и Дополнительные (Additional). Чем больше вы расскажете о своем товаре, тем выше вероятность, что покупатель найдет его на eBay или внешних ресурсах, таких как Google Shopping.
6. В разделе Описание товара (Item description) опишите свой товар в соответствующем поле. Хорошее описание должно быть кратким и структурированным, чтобы покупатель не пропустил важную информацию при беглом просмотре. При этом товар нужно описать достаточно подробно, чтобы не возникло недопонимания: обязательно укажите ключевые характеристики и комплектность. В конце описания добавьте, что к вам можно обращаться с любыми вопросам по поводу товара, и вы обязательно ответите на них. Готовность продавца проконсультировать покупателя и взаимодействовать с ним повышает доверие, а значит и ваш шанс на успешную продажу.
Помните, что именно описание в вашем объявлении — это первая подробная информация о вашем товаре, которую видит покупатель. Здесь вы можете написать все, что сочтете нужным и важным для него.
Примечание: в целях защиты покупателей eBay ограничивает ссылки (в том числе нерабочие) на сторонние сайты. Допускаются ссылки на видео о продукте, другие страницы eBay, сайты ряда транспортных компаний и сторонних поставщиков, а также на необходимую по закону информацию. Более подробная информация доступна здесь (на английском языке).
Более подробная информация доступна здесь (на английском языке).
В правом верхнем углу Карточки создания объявления расположены две кнопки: Проверка на совместимость с мобильными устройствами (Mobile-friendly checker) и Расширенное редактирование (Advanced editing).
Первая кнопка покажет вам, как текст описания будет выглядеть на экране смартфона. Возможно, его стоит сократить или разделить на короткие параграфы для удобства чтения.
Расширенное редактирование позволит вам менять шрифт, размер, цвет текста, а также настраивать отступы и выравнивание.
Примечание: вы можете переключиться на вкладку HTML, чтобы получить больше возможностей для редактирования и поисковой оптимизации. Но для работы с HTML-редактором вам понадобятся специальные навыки.
Определите Условия продажи (Selling details)
Один из важнейших этапов создания объявления — определение условий продажи. Для этого заполните поля соответствующего раздела.
1. В поле Формат (Format) выберите тип торгов: Аукцион (Auction-style) или Фиксированная цена (Fixed price).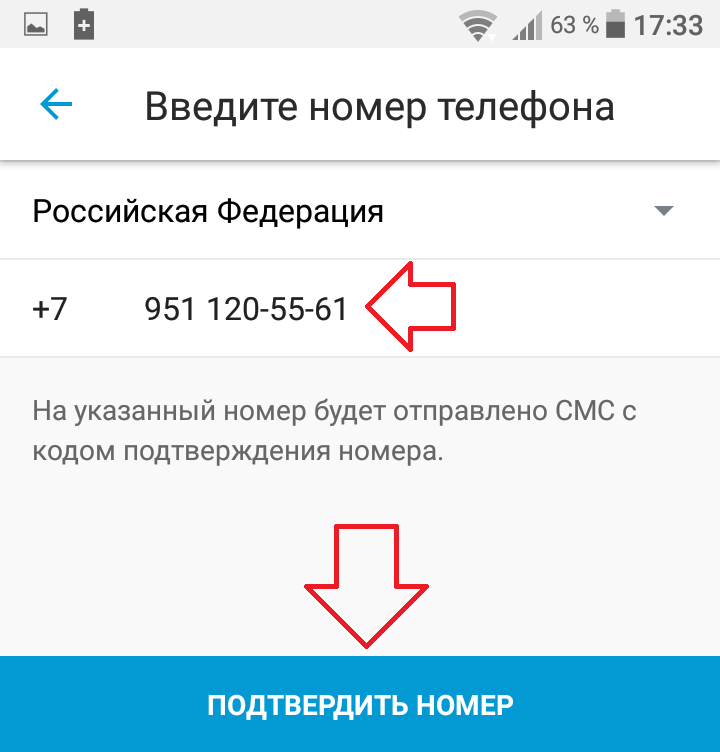
2. В поле Продолжительность (Duration) определите срок действия объявления: от 3 до 10 дней или Годен до отмены (Good ‘Til Cancelled). Здесь же вы можете выбрать время публикации вашего объявления: сразу же после окончания редактирования (Start my listings when I submit them) или в определенный день и время (Schedule to start on).
3. В поле Цена (Price) задайте стоимость товара. Если вы размещаете объявление с фиксированной ценой (Fixed price), укажите цену Купить сейчас (Buy It Now price). Для формата Аукцион (Auction-style) укажите начальную (Starting price) и цену Купить сейчас (Buy It Now price). За дополнительную плату можно установить резервную цену (Reserve price) — минимальную цену, которую вы хотите получить за товар в ходе аукциона. Резервная цена будет скрыта от покупателей.
Примечание: если вы хотите увеличить шансы на успешную продажу, предоставьте покупателям возможность предлагать свою цену. Для этого подключите опцию Лучшее предложение (Best offer).
4. В поле Количество (Quantity) укажите количество доступного товара. Если вы хотите продать коллекцию или несколько товаров одному покупателю, воспользуйтесь опцией Продать набором (Sell as a lot).
4. Опция Частное объявление (Private listing) дает возможность покупателю совершить покупку, сохраняя анонимность.
5. Опция Сделайте благотворительный взнос (Make a donation) позволяет продавцу автоматически переводить часть полученных от продажи деньги в выбранный благотворительный фонд. Если вы подключите эту опцию, eBay снизит ваши базовые сборы с продажи.
6. Вы можете указать размер Налога с продаж (Sales tax), который будет автоматически применяться к цене вашего товара.
7. В настройках возврата отметьте, принимаете ли вы Возвраты в стране местонахождения (Domestic returns) или Международные возвраты (International returns). Также укажите срок возврата и того, кто его оплачивает, — покупателя (Buyer) или продавца (Seller). Рекомендуется принимать возвраты: это увеличивает шансы на успешную продажу товара.
Рекомендуется принимать возвраты: это увеличивает шансы на успешную продажу товара.
Примечание: если вы размещаете объявление на сайте ebay.com, помните, что опция Возвраты в стране местонахождения относится только к территории США. Больше о настройках возврата читайте здесь.
Настройте Условия доставки (Shipping details)
В этом разделе укажите условия доставки и регионы, в которые вы будете доставлять свой товар. Подробную инструкцию и полезные советы ищите в разделе Как настроить доставку.
Примечание: на сайте ebay.com Внутренняя доставка (Domestic shipping) означает доставку в пределах США.
Продайте товар быстрее (Sell it faster)
Вы можете привлечь больше покупателей, если предложите им скидку за покупку нескольких товаров (Volume pricing).
Публикация объявления
Ваше объявление почти готово. Меню внизу страницы поможет вам увидеть, что получилось в итоге. Обратите внимание на поле Сборы (Fees): в нем указана стоимость выставления данного товара.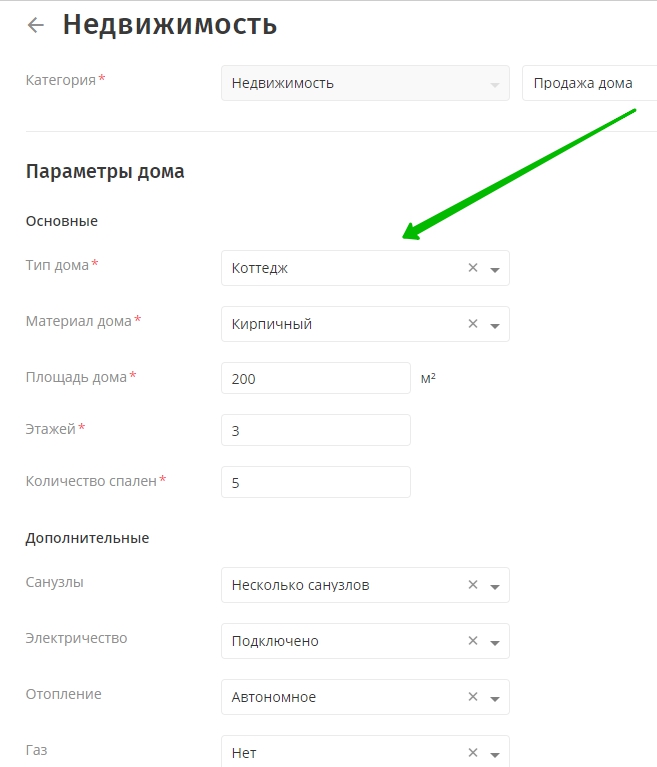 Если в поле задано значение, отличное от нуля, объявление будет платным.
Если в поле задано значение, отличное от нуля, объявление будет платным.
1. Нажав на кнопку Предварительный просмотр (Preview), вы увидите, как будет выглядеть ваше объявление для посетителей eBay.
2. Нажав на кнопку Сохранить как черновик (Save as draft), вы можете сохранить свое объявление для последующего редактирования, без публикации.
3. Нажав на кнопку Отменить (Cancel), вы отмените создание объявления.
Наконец, нажав на кнопку Опубликовать (List item), вы разместите объявление на eBay.
Поздравляем! Ваше первое объявление готово
В дальнейшем вы можете сохранять незаконченные объявления, чтобы закончить работу с ними позже. Раздел Сохраненные черновики (Latest drafts) находится на главной странице Карточки создания объявления под полем ввода. Рядом с иконкой каждого черновика есть две кнопки: Продолжить работу над объявлением (Resume) и Удалить (Delete).
Черновики также можно найти в Центре продавцов: на вкладке Объявления (Listings), в разделе Управление черновиками (Manage drafts).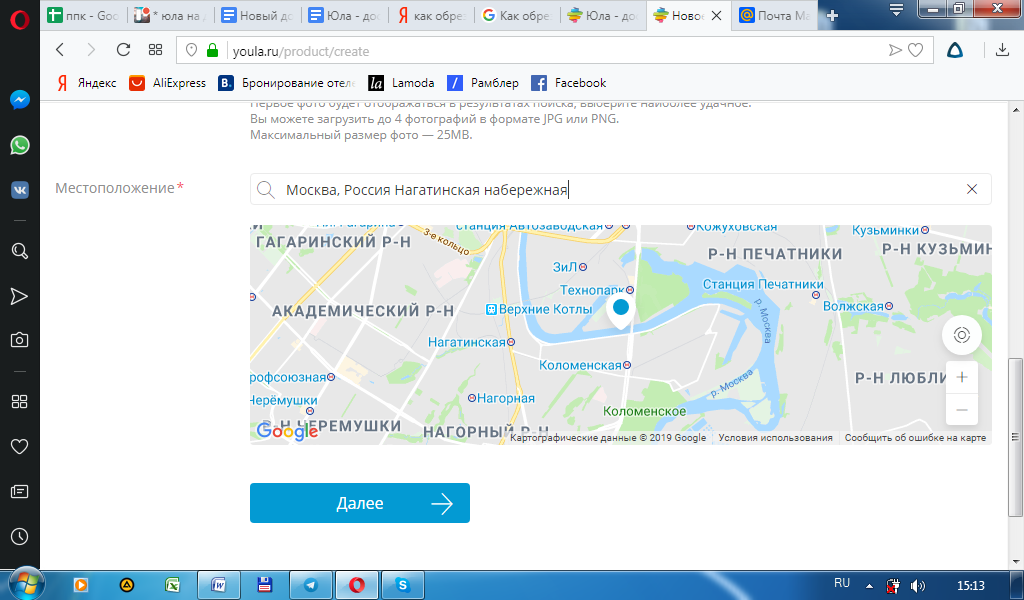
Также на странице Карточки создания объявления есть раздел Шаблоны объявлений (Listing templates). Шаблоны помогают автоматизировать создание объявлений для похожих товаров. Для создания шаблона объявления нажмите на кнопку Создать шаблон (Create a template). Вы будете автоматически перенаправлены в Карточку шаблона. Она практически идентична Карточке создания объявления и заполняется аналогичным образом. Позже вы сможете использовать сохраненный шаблон для быстрого создания новых объявлений.
Желаем вам успешных продаж!
Черпайте вдохновение в историях успеха наших продавцов.
В случае любых затруднений обращайтесь в Службу поддержки.
Часто задаваемые вопросы
Для начинающих продавцов на ebay.com действует лимит: в течение месяца можно выставить не более 10 товаров на общую сумму не более 500 долларов США. При этом учитывается каждая единица товара независимо от количества объявлений. Ограничения разработаны, чтобы помочь новичкам освоиться на площадке, не делать большого количества ошибок, планово развивать бизнес и налаживать отличный клиентский сервис.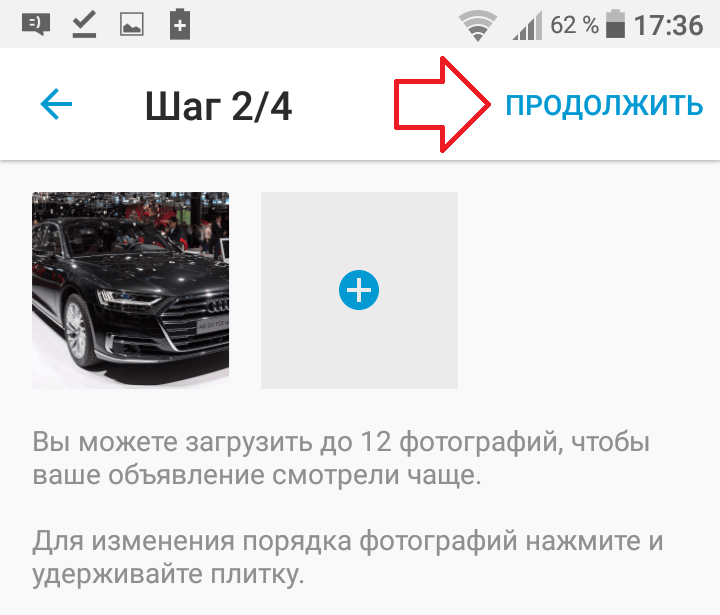
eBay будет ежемесячно проверять вашу учетную запись и автоматически корректировать лимиты в зависимости от объема продаж и отзывов, полученных от покупателей. Вы всегда можете посмотреть действующие лимиты во вкладке Обзор в Центре продавцов. Для этого прокрутите страницу вниз и найдите раздел Ежемесячные лимиты. Там же расположена ссылка для запроса на размещение большего количества товаров.
Кроме лимитов, которые касаются общего объема продаж, на ebay.com существуют ограничения в рамках отдельных категорий товаров.
Каждый месяц вы можете разместить на ebay.com до 250 бесплатных объявлений, то есть без Сбора за выставление товара. После того как ежемесячная Квота будет исчерпана, за выставление каждого объявления в большинстве категорий нужно будет заплатить 0,35 долларов США.
Для подписчиков на Магазин eBay Квота на бесплатные объявления и скидки на Сбор за выставление увеличиваются в зависимости от уровня подписки. Размеры Квоты и Сбора за выставление товара также могут зависеть от стоимости и категории товара, правил регионального сайта eBay (например, ebay. de, ebay.it и др.), а также от типа вашей учетной записи — является ли она личной (Private Seller) или компании (Business Seller).
de, ebay.it и др.), а также от типа вашей учетной записи — является ли она личной (Private Seller) или компании (Business Seller).
На eBay выставляется около 1,5 миллиардов объявлений от миллионов продавцов из разных стран мира. На площадке можно увидеть практически любые товары — новые и подержанные, фабричного производства и ручной работы, массовые и уникальные, разных категорий и ценовых уровней.
При этом eBay серьезно относится к безопасности сообщества, поэтому некоторые виды и категории товаров запрещено выставлять на продажу, либо можно выставить с рядом ограничений. Кроме того, перед размещением объявления убедитесь, что вы не нарушаете местные законы тех стран, в которые собираетесь продавать товар.
Если вы еще не определились с ассортиментом продаж, изучите, какие товары популярны на eBay в целом и у продавцов из вашего региона, чтобы предлагать то, что будет востребовано.
Информация была полезна?
WORKMAN привлекает потенциальных клиентов к рекламе в рождественском журнале.

По данным Ассоциации очагов, патио и барбекю, только около половины (52 процента) американских домов имеют камин. Для тех 48 процентов из нас, у кого нет дымохода, нелегко проникнуться праздничным духом, особенно когда почти все популярные представления о Рождестве вращаются вокруг каштанов, жарящихся на открытом огне, и Санта-Клауса, спускающегося по дымоходу.
К счастью, есть замена: Святочное полено.
Скорее всего, вы уже заметили и, вероятно, смотрели один из этих суррогатных праздничных каминов. Но на случай, если вы этого не сделали, йольский журнал выглядит так: это видео камина с горящими внутри поленьями. Вы смотрите это. Вот и все.
Некоторые журналы Йола могут быть довольно сложными. Виртуальный камин, например, позволяет зрителям выбирать свою собственную стопку поленьев и даже выбирать из нескольких разных звуков горения. Некоторые из них носят тематический характер, например Рождественский журнал Дарта Вейдера, в котором на бревнах изображены останки знаменитого злодея. (Говорите, что хотите, но у этого материала более 1,2 миллиона просмотров.) В третьих есть интернет-звезды. У знаменитого кота Лил Баба, например, есть собственное видео из дневника Рождества.
(Говорите, что хотите, но у этого материала более 1,2 миллиона просмотров.) В третьих есть интернет-звезды. У знаменитого кота Лил Баба, например, есть собственное видео из дневника Рождества.
«В реальной жизни в огне есть что-то волшебное, и по какой-то причине он все еще работает на уровне экрана», — объясняет основатель Virtual Fireplace Йерун Клэп. «Даже люди, которые поначалу настроены скептически, обнаружат, что они автоматически продолжают смотреть на огонь на экране и становятся спокойными и уютными. Каким-то образом вид контролируемого огня, реального или на экране, дает нам чувство безопасности и благополучия. -существование.»
Брендинг у камина
Но, как обнаружили многие бренды, вид контролируемого огня на экране также может быть хорошим маркетинговым ходом. В журнале Happy Yule Log, дебютировавшем в прошлом году, были изображены рыжий кот по имени Хэппи и собака (также по имени Хэппи), и он был предоставлен вам каналом Hallmark. (Хэппи и Хэппи, оба спасенные из приютов, также снялись в новом видео из журнала Йола 2016 года, трехчасовой трансляции, премьера которой состоялась в День Благодарения и которая вернется в канун Рождества. )
)
В 2015 году Budweiser и односолодовый виски Lagavulin разместили фирменные видеоролики Yule Log, в первом из которых Clydesdale жует сено у очага (см.
В этом году в огонь попало несколько других брендов. Hearthstone — фэнтезийная карточная онлайн-игра, принадлежащая Blizzard Entertainment, — выложила дневник Рождества, в котором изображены его фирменные существа (называемые мурлоками), пытающиеся (хотя и не совсем успешно) петь праздничную музыку, пока горят поленья.
«В то время как рождественские журналы становятся популярными среди брендов, Hearthstone находится в уникальном положении, потому что очаг всегда был такой важной частью опыта», — сказал представитель Blizzard Entertainment. «Здесь наши игроки приходят на связь, независимо от того, входят ли они в таверну Hearthstone со своих телефонов или компьютеров или посещают местное мероприятие Fireside Gathering».
Представитель Blizzard воздержался от того, чтобы назвать йольский журнал маркетинговым ходом, но сказал, что «мы хотим сделать опыт [игрового сообщества] как можно более эпическим», и видео является частью этого. Она добавила, что некоторые поклонники Hearthstone действительно тратят целый час на просмотр видео из журнала «Рождество», которое на момент публикации набрало около 728 000 просмотров.
Она добавила, что некоторые поклонники Hearthstone действительно тратят целый час на просмотр видео из журнала «Рождество», которое на момент публикации набрало около 728 000 просмотров.
Затем идет йольский журнал для Xbox игры Dead Rising 4. Игра, сюжет которой уже разворачивается во время праздников, обещает «массовое уничтожение зомби», и видео из йольского журнала это доказывает. Под веселый напев праздничной мелодии главный герой Фрэнк Уэст подходит к камину, чтобы периодически подбрасывать руки и ноги убитых им зомби в огонь, где они потрескивают и трещат. Так что, да, это не то видео, которое можно поставить бабушке.
«Франшиза Dead Rising известна тем, что она превосходна и никогда не боится раздвигать границы», — объяснил руководитель отдела маркетинга игры Генри Лю. «Мы построили маркетинговую кампанию на идее «захвата праздников» — мы взяли традиционные праздничные образы, но придали им оттенок Dead Rising».
«Каждый знаком с видео из Святочного журнала — праздничной традицией, простой, но веселой для всех. Включив зомби в уравнение, мы смогли создать что-то, что отличалось от других видео из Святочного журнала, но полностью соответствовало духу Мертвых. Растущий бренд». (XBox позаботился о том, чтобы поместить предупреждение о «крови и увечьях» прямо перед тем, как начнется воспроизведение видео.)
Включив зомби в уравнение, мы смогли создать что-то, что отличалось от других видео из Святочного журнала, но полностью соответствовало духу Мертвых. Растущий бренд». (XBox позаботился о том, чтобы поместить предупреждение о «крови и увечьях» прямо перед тем, как начнется воспроизведение видео.)
Пламя может разгореться сильнее
Несмотря на то, что такие гиганты, как Anheuser-Busch InBev и Microsoft, зажгли рождественские бревна в качестве идея брендинга, некоторые считают, что эта идея остается недостаточно развитой.
«Я всегда думал, что есть способы монетизировать их с помощью брендинга», — отметил Петур Воркман, генеральный директор Workman Global, , бренд-агентства, которое налаживает отношения между креативными компаниями и средними и крупными брендами. «Это не было бы чем-то вопиющим — не «купи мой продукт», — но очаг или каминная полка могли бы содержать что-то более подсознательное».
Например, Уоркман предположил, что, если бы над камином была рамка для картины, которая могла бы содержать сообщение бренда? Или что, если бы вы могли убедить музыкального исполнителя, что его или ее песни будут играть фоном? Журналы Digital Yule, по словам Воркмана, «великолепны. Я думаю, что они гениальны. Но я не вижу, чтобы они максимально использовали свои возможности».
Я думаю, что они гениальны. Но я не вижу, чтобы они максимально использовали свои возможности».
Вы можете спросить об этом Клапа. Когда он запустил Virtual Fireplace в 2014 году, это было просто хобби, то есть до тех пор, пока Google не предложил ему присоединиться к программе AdSense, и он обнаружил, что может зарабатывать деньги, предоставляя свои цифровые журналы Yule различным компаниям, которые, в свою очередь, используют их для маркетинг. Доход от его первых святочных журналов теперь позволяет ему создавать новые.
«Я продаю коммерческие лицензии самым разным компаниям, от домов престарелых, в которых не могут быть настоящие камины из соображений безопасности, до компаний кабельного телевидения, которые хотят предоставить своим подписчикам расслабляющий каминный канал», — сказал Клэп.
На самом деле, спустя два года после того, как Клап начал свой праздничный бизнес, большая проблема заключается не в том, чтобы продать огонь, а в том, чтобы найти камины для стрельбы (это один из них выше), поскольку у него нет своего собственного. «Я всегда ищу людей, у которых есть хороший камин, и спрашиваю их, могу ли я его снять. Иногда люди смотрят через мое плечо, чтобы убедиться, что я не сожгу их дом».
«Я всегда ищу людей, у которых есть хороший камин, и спрашиваю их, могу ли я его снять. Иногда люди смотрят через мое плечо, чтобы убедиться, что я не сожгу их дом».
Нравятся эти журналы? Спасибо WPIX
За сегодняшней кучей бревен скрывается еще один животрепещущий вопрос: как вообще возникла идея наблюдать за камином на экране?
На самом деле все началось ровно 50 лет назад, когда менеджер нью-йоркской телестанции WPIX Фред Троуэр принял судьбоносное решение приостановить программы по расписанию в пользу трех часов горящего камина. С благословения мэра Джона Линдсея съемочная группа PIX сняла кадры на 16-миллиметровой пленке в особняке Грейси, официальной резиденции мэра. Техники соединили две минуты фильма вместе, чтобы получить три часа Рождественского журнала, добавили к нему немного рождественской музыки и выпустили в эфир в 9 часов.:30 вечера В канун рождества. «Это должно было стать рождественской открыткой для жителей Нью-Йорка», — сказал директор WPIX по цифровой и социальной стратегии Роландо Пухоль. «Это никогда не должно было быть чем-то, на чем мы могли бы заработать». (На самом деле, упреждая запланированную программу — роллер-дерби, в первую очередь — станция потеряла 4000 долларов в тот первый год. Это почти 30 000 долларов в сегодняшних долларах.) , завоевав своего рода добрую волю среди зрителей, которая имела собственную ценность, хотя, возможно, и не поддающуюся количественной оценке.
«Это никогда не должно было быть чем-то, на чем мы могли бы заработать». (На самом деле, упреждая запланированную программу — роллер-дерби, в первую очередь — станция потеряла 4000 долларов в тот первый год. Это почти 30 000 долларов в сегодняшних долларах.) , завоевав своего рода добрую волю среди зрителей, которая имела собственную ценность, хотя, возможно, и не поддающуюся количественной оценке.
Пухоль работает в WPIX только с 2012 года, но в этом году он оказался маловероятным игроком в длинной истории рождественского журнала. Оригинальные кадры 1966 года пропали где-то после 1969 года, когда пленка износилась, и PIX решили вместо этого снять другой камин. (Поскольку во время оригинальной съемки искра из камина сожгла дорогой ковер, мэр не пригласил PIX обратно в особняк Грейси, и вместо этого PIX нашел новый камин в доме в Калифорнии.)
Оригинальные кадры считались утерянными. навсегда, пока в 2014 году Пужоль не позвонила женщина из Парамуса, штат Нью-Джерси. Она продавала дом, который когда-то принадлежал покойному Биллу Куперу, давнему руководителю отдела фильмов и кассет WPIX. Звонивший сообщил Пухолю, что у нее есть гараж, полный старых канистр от пленки, и спросил, не хочет ли он их получить. «Мы загрузили два фургона и привезли все это в подвал», — рассказал Пухоль.
Звонивший сообщил Пухолю, что у нее есть гараж, полный старых канистр от пленки, и спросил, не хочет ли он их получить. «Мы загрузили два фургона и привезли все это в подвал», — рассказал Пухоль.
А теперь перенесемся в это лето. «Сейчас июль 2016 года, и я сижу в подвале и ищу кадры с Дональдом Трампом в 1980-х», — сказал он. «Некоторые банки с пленкой стояли на полке на уровне глаз. Я проходил мимо, и передо мной выскочила банка с надписью: «Ориг. Камин WPIX». был восстановлен, и он будет транслироваться в этом году, начиная с Сочельника.
За исключением нескольких лет в 1990-х годах, когда йольский журнал был в перерыве, WPIX по-прежнему каждое Рождество упреждает несколько часов коммерческих программ, чтобы позволить этим журналам гореть, решение, которое становится все более трудным, поскольку традиционное телевидение изо всех сил пытается оставаться конкурентоспособным. Ранний утренний интервал, который будет транслироваться в журнале Йоля, — это время, когда станция обычно запускала рекламные ролики, «и это наш хлеб с маслом», — сказал Пухоль. Тем не менее, он твердо верит, что часы этого праздничного камина в конечном итоге приносят пользу бренду WPIX.
Тем не менее, он твердо верит, что часы этого праздничного камина в конечном итоге приносят пользу бренду WPIX.
«Когда вы думаете о современном мире с его разнообразием информации, все, что мы можем сделать, чтобы сохранить идентичность WPIX, дать ему ощущение места, сейчас важнее, чем это было в 1966 году», — сказал он. «Большая часть того, что мы показываем в эфире, является товарным контентом. Но журнал Йоля уникален и особенный. Он оказывает эффект ореола на все остальное, что мы делаем».
Для получения полной информации см. «Как Yule Logs превратился из причудливого в популярный маркетинговый инструмент» Роберта Клары — 8 декабря 2016 г.
Yule Love This Beautiful Ads (ВИДЕО)
Posten Norge, Disney UK, George at Asda, Boots
В этом году порция рождественских рекламных роликов только что вышла из духовки, а рекламные агентства по обе стороны Атлантики готовят угощения для всех нас в этот праздничный сезон. И мы действительно имеем в виду «всех нас» — во многих телевизионных роликах в этот рождественский праздник особое внимание уделяется инклюзивности.
От однополого романа у камина до нового типа «праздничного бала» — вот реклама, которая нам нравится в этом году.
Posten Norge, «Когда Гарри встретил Санту»
Почтовая служба Норвегии завоевывает поклонников по всему миру этим 4-минутным короткометражным фильмом, в котором рассказывается о нежном романе между одиноким мужчиной и Человеком в красном. Реклама посвящена 50-летию декриминализации гомосексуализма в стране, и Posten Norge сообщила NBC News , что ее цель состояла в том, чтобы «показать, что любовь не имеет границ» и что реклама до сих пор была получена с «ошеломляющим» откликом в Норвегии. .
Etsy, «Наш Санта»
Говоря о лучшем представлении, 30-секундный ролик Etsy показывает, как Санта-Клаус изображался как белый человек с незапамятных времен. Итак, с помощью некой онлайн-площадки для поделок внук делает дедушке подарок, который переворачивает эту традицию.
Итак, с помощью некой онлайн-площадки для поделок внук делает дедушке подарок, который переворачивает эту традицию.
Джон Льюис, «Нежданный гость»
Действительно, нежданный гость! Джон Льюис, британская сеть универмагов, всегда представляет кинематографическую рождественскую рекламу, и эта не является исключением, показывая подростка, знакомящего инопланетянина с традициями Рождества. «Мы знаем, что наши покупатели как никогда взволнованы Рождеством, жаждут радостных моментов, единения и бегства от действительности, поскольку многие из них не смогли отпраздновать с близкими в прошлом году», — пояснил ритейлер в своем заявлении.
TK Maxx, «Christmas to the Maxx»
Здесь, в Штатах, мы получаем рекламу TJ Maxx, призывающую нас стать «максинистами», что бы это ни значило. Но через океан британский коллега универмага представил эту историю о школьном концерте, который получил толчок хард-рокового праздничного духа благодаря паре мерцающих ботинок одного подростка.
Но через океан британский коллега универмага представил эту историю о школьном концерте, который получил толчок хард-рокового праздничного духа благодаря паре мерцающих ботинок одного подростка.
Сапоги, «Сумки радости»
Дженна Коулман из Змей и Виктория звезды славы в этом рекламном ролике, снятом оскароносным режиссером Томом Хупером (который, возможно, пытается искупить свою экранизацию Кошки ). В трехминутном рекламном ролике персонаж Коулман получает волшебную сумочку от своей «Нэн» через британского ритейлера Boots и находит внутри идеальный подарок на любой случай.
Amazon, «Доброта, величайший дар»
Мегакорпорация электронной розничной торговли напоминает нам, что доброта — это величайший дар — конечно, если она приходит в форме покупки на Amazon. А если серьезно, то этот 2,5-минутный фильм, поставленный на новую песню Адель «Держись», рассказывает трогательную историю о женщине, которая успокаивает свою младшую соседку в эпоху пандемии. «Последние 18 месяцев были непростыми для людей во всем мире, в том числе для многих молодых людей. Наше время вместе нельзя воспринимать как должное», — заявил в своем заявлении Эд Смит, генеральный директор по интегрированному маркетингу Amazon в ЕС. «Поэтому в этом году, хотя мир еще не полностью вернется к нормальной жизни, возможности для доброты и общения приобретут новое значение».
А если серьезно, то этот 2,5-минутный фильм, поставленный на новую песню Адель «Держись», рассказывает трогательную историю о женщине, которая успокаивает свою младшую соседку в эпоху пандемии. «Последние 18 месяцев были непростыми для людей во всем мире, в том числе для многих молодых людей. Наше время вместе нельзя воспринимать как должное», — заявил в своем заявлении Эд Смит, генеральный директор по интегрированному маркетингу Amazon в ЕС. «Поэтому в этом году, хотя мир еще не полностью вернется к нормальной жизни, возможности для доброты и общения приобретут новое значение».
Disney UK, «Отчим»
Помните прошлогоднюю рождественскую рекламу Disney UK? Главная героиня этой рекламы, Николь, возвращается в этом новом ролике, в котором основное внимание уделяется ее новому мужу, который переезжает к Рождеству и налаживает связь с ее детьми — конечно, с небольшой помощью персонажей Диснея.
Джордж из Асды, «Slay It»
Категория: Рождество в бальном зале! В этом рекламном ролике для Джорджа, линии одежды в британской сети супермаркетов Asda, исполнитель Джабарии делает скучный паб en vogue на праздники. Более того, Джордж из Asda также пожертвовал 100 000 фунтов стерлингов Diversity Role Models, своему партнеру по рекламе, чтобы помочь положить конец издевательствам над молодежью ЛГБТК+. Мы ставим этому сотрудничеству 10, 10, 10 баллов по всем направлениям!
Лучшие видео Yule Log: YouTube, Netflix, Roku и 4K тоже
Перейти к основному содержаниюПутеводитель Тома поддерживается своей аудиторией. Когда вы покупаете по ссылкам на нашем сайте, мы можем получать партнерскую комиссию. Вот почему вы можете доверять нам.
(Изображение предоставлено: Майкл Бубле/YouTube) Зная, что люди будут искать в Интернете лучшие видеоролики «Рождественского журнала» — в то время как кусачки теряют доступ к традиционным телевизионным трансляциям сжигания поленьев на открытом огне, — я провел работу по поиску такого контента в трех самых популярных местах. Йольские журналы, которые вы должны смотреть, и те, которые вы должны игнорировать.
Йольские журналы, которые вы должны смотреть, и те, которые вы должны игнорировать.
В процессе я понял, что лучшими вариантами являются YouTube и Netflix, хотя первый, по общему признанию, не является идеальным выбором для большинства. Если у вас нет подписки YouTube Premium, ваши часы потоковой передачи Yule Log будут прерываться рекламой.
- Лучшие потоковые устройства
- Лучший Netflix VPN позволит получить еще больше от вашей подписки
- Хотите больше разнообразия просмотра? Вот как изменить регион на Netflix
Многие из лучших потоковых сервисов, таких как Amazon Prime Video, также имеют свои собственные каналы журнала Yule. Apple TV Plus также входит в число тех, где вы можете легко найти журнал Йоля.
Так что приготовь какао и каштаны, дорогой читатель, потому что я нашел лучшие видео Йольского журнала для твоего праздничного сезона. Есть даже один от Майкла Бубле.
Лучшее видео «Святочное полено» на YouTube
Если вы хотите посмотреть видео «Святочное полено», в котором хорошо показаны горящие поленья в камине и звуки, которые они издают, то это видео от Virtual Fireplace поможет вам.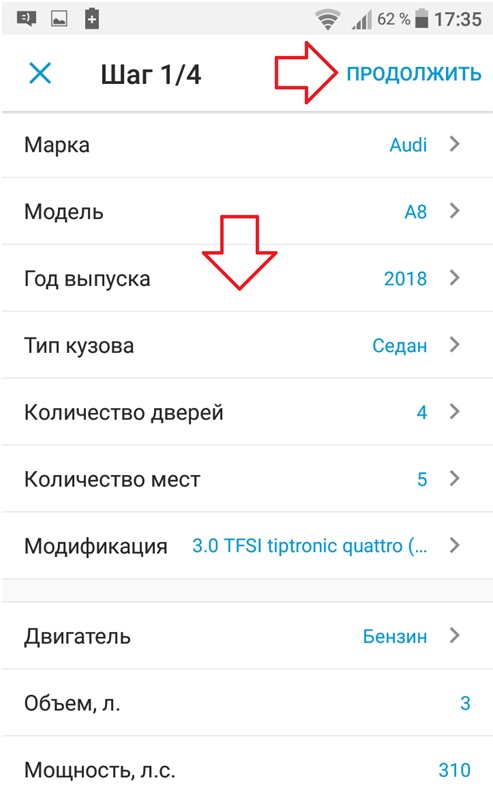 Вероятно, у него самые лучшие и легко слышимые потрескивающие звуки, поэтому вам не нужно увеличивать громкость телевизора до максимума, что приводит к грубому пробуждению, если вы переключаете канал.
Вероятно, у него самые лучшие и легко слышимые потрескивающие звуки, поэтому вам не нужно увеличивать громкость телевизора до максимума, что приводит к грубому пробуждению, если вы переключаете канал.
Единственным недостатком этого видео является максимальное разрешение 720p.
Продолжительность: 2 часа
Если вас больше интересуют визуальные эффекты, а не звуки, это видео, в котором яркое (больше оранжевого и желтого) пламя, вероятно, будет вашим лучшим выбором. Мало того, что это пламя выглядит великолепно, оно также транслируется в разрешении до 1080p, так что журналы чистые. Однако по сравнению с нашим первым выбором его звуковые эффекты не так выражены.
Продолжительность: 10 часов
Что, если вам нужно что-то еще? У Hallmark есть рождественское бревно с кошкой и собакой, которое подойдет либо тем, у кого нет собственного питомца, либо тем, кому было бы смешно, если бы их собственные питомцы пытались взаимодействовать с окружающими. экранные животные.
экранные животные.
Продолжительность: 3 часа
Если вам нужно изображение высочайшего качества, но вы предпочитаете праздничную музыку, а не треск и щелчки, Michael Buble поможет вам. В этом видео «Рождественский бревно» великолепное рождественское бревно в разрешении 4K сочетается с 19 праздничными песнями эстрадного певца. Песни включают «Белое Рождество» (дуэт с Шанайей Твейн) и классику, в том числе «Санта-Клаус едет в город» и «Счастливого Рождества».
Продолжительность: 1 час 5 минут
Лучшие видео из рождественского журнала на Netflix
Netflix предлагает три предложения Yule Log от одного и того же бренда: Fireplace For Your Home. Вот разбивка того, чем они отличаются
Камин для вашего дома (открывается в новой вкладке) состоит из трех «эпизодов», и первый дает вам традиционные Святочные поленья, горящие под праздничную рождественскую музыку на заднем плане. Эпизод 2 заменяет Святочные бревна дровами, а эпизод 3 — это, по сути, первый эпизод с деноминационным саундтреком.
Затем на Netflix есть еще два варианта, и оба в формате 4K UHD: «Камин для вашего дома: классическое издание» (открывается в новой вкладке) и «Камин для вашего дома» Birchwood Edition (открывается в новой вкладке). Они отличаются только поджигаемой древесиной (классика — обычная древесина). Ни в том, ни в другом нет музыкального саундтрека, но я лично выбрал их для лучших видеороликов «Рождественского журнала».
Продолжительность: 1 час каждый
Лучшие видео из журнала Yule Log на Roku
Ненавижу это говорить, но сказать «лучшие видео из журнала Yule Log на Roku» все равно, что сказать «самый ужасный кусок пиццы». По сравнению с тем, что я нашел на YouTube и Netflix, бесплатные варианты Roku интересны только из-за отсутствия ценника.
Лучший из них даже не от Року, а тот, который я нашел в поиске Року. Это The RiffTrax YuleLog (откроется в новой вкладке) (2 часа), который доступен бесплатно на Tubi. Его визуальные эффекты в порядке, и есть некоторые словесные риффы, как и в случае с RiffTrax.
Его визуальные эффекты в порядке, и есть некоторые словесные риффы, как и в случае с RiffTrax.
Затем есть предложение от Roku Yule Log Channel (открывается в новой вкладке) (15,5 часов), которое настолько низкое, что я подумал, что оно должно воссоздавать то, что это было все равно, что смотреть оригинальную трансляцию Йольского журнала. Глядя на мелкий шрифт, кажется, что это не так.
Если вам нравится Святочное полено с привкусом сахарина, есть кое-что под названием Официальный праздничный Святочный камин (открывается в новой вкладке) (30 минут), который украшен фестонами. Если бы пылающий лес был более заметен, я был бы более позитивен, но это очень мало для Йольского бревна.
Лучшие на сегодняшний день предложения Amazon по продаже 55-дюймовых 4K-телевизоров
804 Отзывы покупателей Amazon (открывается в новой вкладке)
☆☆☆☆☆
(открывается в новой вкладке)
(открывается в новой вкладке)
0002 $889,99(открывается в новой вкладке)
Просмотр сделки (открывается в новой вкладке)
Низкий запас
(открывается в новой вкладке)
(открывается в новой вкладке)
(открывается в новой вкладке)
$899
(откроется в новой вкладке)
Просмотреть предложение (откроется в новой вкладке)
Цена со скидкой
(откроется в новой вкладке)
(откроется в новой вкладке)
1999,99 $ новая вкладка (откроется в новой вкладке)
90 дюймов1699 долларов
(открывается в новой вкладке)
Посмотреть предложение (откроется в новой вкладке)
Показать больше предложений
Генри — старший редактор Tom’s Guide, освещающий потоковое мультимедиа, ноутбуки и все, что связано с Apple, обзор устройств и сервисов за последние семь лет. До прихода в Tom’s Guide он рецензировал программное и аппаратное обеспечение для TechRadar Pro и брал интервью у художников для международного журнала Patek Philippe. Он также освещал дикий мир профессионального рестлинга для Cageside Seats, брал интервью у спортсменов и других ветеранов индустрии.
До прихода в Tom’s Guide он рецензировал программное и аппаратное обеспечение для TechRadar Pro и брал интервью у художников для международного журнала Patek Philippe. Он также освещал дикий мир профессионального рестлинга для Cageside Seats, брал интервью у спортсменов и других ветеранов индустрии.
Темы
Потоковое
Tom’s Guide является частью Future US Inc, международной медиагруппы и ведущего цифрового издателя. Посетите наш корпоративный сайт (откроется в новой вкладке).
© Future US, Inc. Полный 7-й этаж, 130 West 42nd Street, Нью-Йорк, Нью-Йорк 10036.
Разграничение видов с использованием данных по одному локусу и обобщенного подхода к слиянию смешанного юла: пересмотренный метод и оценка смоделированных наборов данных | Систематическая биология
Журнальная статья
Томочика Фудзисава,
Томочика Фудзисава
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Тимоти Г. Барраклаф
Барраклаф
Тимоти Дж. Барраклаф *
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Примечания автора
Систематическая биология , том 62, выпуск 5, сентябрь 2013 г., страницы 707–724, https://doi.org/10.1093/sysbio/syt033
Опубликовано:
3 июнь 223
1 История статьи
Получено:
11 августа 2012 г.
Получена редакция:
19 ноября 2012 г.
Принято:
03 мая 2013 г.
Опубликовано:
14 июня 2013 г.
- Разделенный вид
- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Cite
Tomochika Fujisawa, Timothy G.
 Barraclough, Разграничение видов с использованием данных по одному локусу и обобщенного смешанного коалесцентного подхода: пересмотренный метод и оценка смоделированных наборов данных, Систематическая биология , том 62, выпуск 5, сентябрь 2013 г., страницы 707–724, https://doi.org/10.1093/sysbio/syt033
Barraclough, Разграничение видов с использованием данных по одному локусу и обобщенного смешанного коалесцентного подхода: пересмотренный метод и оценка смоделированных наборов данных, Систематическая биология , том 62, выпуск 5, сентябрь 2013 г., страницы 707–724, https://doi.org/10.1093/sysbio/syt033Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Фейсбук
- Еще
 Barraclough, Разграничение видов с использованием данных по одному локусу и обобщенного смешанного коалесцентного подхода: пересмотренный метод и оценка смоделированных наборов данных, Систематическая биология , том 62, выпуск 5, сентябрь 2013 г., страницы 707–724, https://doi.org/10.1093/sysbio/syt033
Barraclough, Разграничение видов с использованием данных по одному локусу и обобщенного смешанного коалесцентного подхода: пересмотренный метод и оценка смоделированных наборов данных, Систематическая биология , том 62, выпуск 5, сентябрь 2013 г., страницы 707–724, https://doi.org/10.1093/sysbio/syt033Фильтр поиска панели навигации Систематическая биологияЭтот выпускЭволюционная биологияКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Систематическая биологияЭтот выпускЭволюционная биологияКнигиЖурналыOxford Academic Термин поиска на микросайте
Advanced Search
Abstract
Исследования типа ДНК-штрихкодирования собирают данные по одному локусу из больших выборок особей и видов и предоставляют новые виды данных для эволюционных исследований разнообразия. Важной целью многих таких исследований является разграничение эволюционно значимых видовых единиц, особенно в исследованиях биоразнообразия на основе образцов ДНК из окружающей среды. Метод Generalized Mixed Yule Coalescent (GMYC) представляет собой вероятностный метод разграничения видов путем подгонки внутривидовых и межвидовых моделей ветвления к реконструированным генным деревьям. Хотя этот метод широко использовался, ранее он не был подробно описан или полностью оценен в сравнении с моделированием альтернативных сценариев истинных закономерностей изменчивости популяций и дивергенции между видами. Здесь мы представляем важные переформулировки метода GMYC в первоначальном виде и демонстрируем его устойчивость к целому ряду отклонений от его упрощающих предположений. Основным фактором, влияющим на точность разграничения, является средний размер популяции видов по отношению к времени дивергенции между ними. Другие отклонения от допущений модели, такие как разный размер популяции видов, альтернативные сценарии видообразования и вымирания, а также рост или подразделение популяции внутри вида, имеют относительно меньшие последствия.
Важной целью многих таких исследований является разграничение эволюционно значимых видовых единиц, особенно в исследованиях биоразнообразия на основе образцов ДНК из окружающей среды. Метод Generalized Mixed Yule Coalescent (GMYC) представляет собой вероятностный метод разграничения видов путем подгонки внутривидовых и межвидовых моделей ветвления к реконструированным генным деревьям. Хотя этот метод широко использовался, ранее он не был подробно описан или полностью оценен в сравнении с моделированием альтернативных сценариев истинных закономерностей изменчивости популяций и дивергенции между видами. Здесь мы представляем важные переформулировки метода GMYC в первоначальном виде и демонстрируем его устойчивость к целому ряду отклонений от его упрощающих предположений. Основным фактором, влияющим на точность разграничения, является средний размер популяции видов по отношению к времени дивергенции между ними. Другие отклонения от допущений модели, такие как разный размер популяции видов, альтернативные сценарии видообразования и вымирания, а также рост или подразделение популяции внутри вида, имеют относительно меньшие последствия. Наши симуляции демонстрируют, что меры поддержки, полученные из функции правдоподобия, обеспечивают надежную индикацию того, когда модель работает хорошо, а когда она приводит к неточным разграничениям. Наконец, так называемая однопороговая версия метода превосходит многопороговую версию метода на смоделированных данных: мы утверждаем, что это может представлять собой фундаментальное ограничение из-за характера доказательств, используемых для разграничения видов в этом подходе. Вместе с другими исследованиями, сравнивающими его эффективность с другими методами, наши результаты подтверждают надежность GMYC как инструмента для разграничения видов, когда доступна информация только об одном локусе. [Кластеры; коалесцирующий; ДНК; генеалогический; нейтральный; видообразование; виды.]
Наши симуляции демонстрируют, что меры поддержки, полученные из функции правдоподобия, обеспечивают надежную индикацию того, когда модель работает хорошо, а когда она приводит к неточным разграничениям. Наконец, так называемая однопороговая версия метода превосходит многопороговую версию метода на смоделированных данных: мы утверждаем, что это может представлять собой фундаментальное ограничение из-за характера доказательств, используемых для разграничения видов в этом подходе. Вместе с другими исследованиями, сравнивающими его эффективность с другими методами, наши результаты подтверждают надежность GMYC как инструмента для разграничения видов, когда доступна информация только об одном локусе. [Кластеры; коалесцирующий; ДНК; генеалогический; нейтральный; видообразование; виды.]
Фундаментальная закономерность природы заключается в том, что организмы диверсифицируются в более или менее дискретные сущности, которые мы называем видами. Термин «вид» используется для обозначения многих аспектов модели разнообразия и его основных причин (Hey 2001; Coyne and Orr 2004; De Queiroz 2007; Shaffer and Thomson 2007). Однако в общих эволюционных терминах (например, в соответствии с общей концепцией происхождения видов, De Queiroz 2007) виды представляют собой группы организмов, которые развиваются независимо от других таких групп (из-за барьеров на пути распространения генов от одного вида к другому). ), которые поэтому расходятся на дискретные единицы морфологической и генетической изменчивости, очевидные из обзоров более высоких клад. В исследованиях природы и происхождения видов все чаще используются эмпирические инструменты для разграничения эволюционно значимых таксонов на основе поддающихся измерению величин, например репродуктивной изоляции (Койн и Орр 19).98; Knowles and Carstens 2007), экологическое расхождение (Fontaneto et al. 2007; Leaché et al. 2009) и другие черты (Feulner et al. 2007), вместо того чтобы полагаться на качественные таксономические методы (см. обсуждение Sites and Marshall 2003).
Однако в общих эволюционных терминах (например, в соответствии с общей концепцией происхождения видов, De Queiroz 2007) виды представляют собой группы организмов, которые развиваются независимо от других таких групп (из-за барьеров на пути распространения генов от одного вида к другому). ), которые поэтому расходятся на дискретные единицы морфологической и генетической изменчивости, очевидные из обзоров более высоких клад. В исследованиях природы и происхождения видов все чаще используются эмпирические инструменты для разграничения эволюционно значимых таксонов на основе поддающихся измерению величин, например репродуктивной изоляции (Койн и Орр 19).98; Knowles and Carstens 2007), экологическое расхождение (Fontaneto et al. 2007; Leaché et al. 2009) и другие черты (Feulner et al. 2007), вместо того чтобы полагаться на качественные таксономические методы (см. обсуждение Sites and Marshall 2003).
Генетические деревья представляют собой ценный источник информации для определения моделей и процессов диверсификации. До недавнего времени исследования диверсификации были ограничены компромиссом между количеством локусов, количеством особей на вид и количеством видов, которые можно было отобрать, что приводило либо к наборам данных о многих локусах у нескольких видов, либо о нескольких локусах для нескольких видов. много видов. Популяционно-генетические и филогеографические исследования изучали историю популяций и поток генов в родственных сестринских видах или видовых комплексах (Templeton 2001; Avise 2009).). В этих исследованиях обычно отбиралось большое количество особей каждого вида и часто несколько молекулярных маркеров (Koufopanou et al. 1997), однако из-за уровня выборки, необходимого для вывода о популяционных процессах, данное исследование по необходимости обычно сосредотачивалось на нескольких видах. Напротив, филогенетические исследования реконструировали события видообразования, приведшие к современным видам, и сделали выводы о макроэволюционных процессах в широких таксономических и географических масштабах (Barraclough and Nee 2001). Однако, поскольку филогенетические исследования требуют большой выборки видов для большой клады, по необходимости в этих исследованиях до недавнего времени (Carstens and Dewey 2010; Camargo et al. 2012) в основном отбирали только один образец для каждого таксономического вида. Теория анализа популяции и филогенетических данных также развивалась отдельно.
До недавнего времени исследования диверсификации были ограничены компромиссом между количеством локусов, количеством особей на вид и количеством видов, которые можно было отобрать, что приводило либо к наборам данных о многих локусах у нескольких видов, либо о нескольких локусах для нескольких видов. много видов. Популяционно-генетические и филогеографические исследования изучали историю популяций и поток генов в родственных сестринских видах или видовых комплексах (Templeton 2001; Avise 2009).). В этих исследованиях обычно отбиралось большое количество особей каждого вида и часто несколько молекулярных маркеров (Koufopanou et al. 1997), однако из-за уровня выборки, необходимого для вывода о популяционных процессах, данное исследование по необходимости обычно сосредотачивалось на нескольких видах. Напротив, филогенетические исследования реконструировали события видообразования, приведшие к современным видам, и сделали выводы о макроэволюционных процессах в широких таксономических и географических масштабах (Barraclough and Nee 2001). Однако, поскольку филогенетические исследования требуют большой выборки видов для большой клады, по необходимости в этих исследованиях до недавнего времени (Carstens and Dewey 2010; Camargo et al. 2012) в основном отбирали только один образец для каждого таксономического вида. Теория анализа популяции и филогенетических данных также развивалась отдельно.
Теперь можно отобрать несколько особей большинства видов из более широкой клады. Например, проекты ДНК-штрихкодирования выполняют молекулярную инвентаризацию больших выборок видов и нескольких особей внутри вида (Hebert et al. 2003; Meyer and Paulay 2005; Monaghan et al. 2009). Точно так же образцы окружающей среды некультивируемых организмов, таких как бактерии и микробные эукариоты, могут быть секвенированы из морских и наземных экосистем (Acinas et al. 2004). Эти данные дают возможность разграничить эволюционно значимые единицы разнообразия в широком таксономическом масштабе. Однако в настоящее время крупномасштабные исследования биоразнообразия по-прежнему в основном полагаются на одиночные локусы, такие как цитохромоксидаза I для штрих-кодов животных или 16S рДНК для исследований разнообразия бактерий (обратите внимание, что штрих-кодирование ДНК растений использует несколько локусов, но часто несколько связанных локусов в пластидном геноме). , что соответствует одному локусу с точки зрения генеалогической информации, см. CBOL Plant Working Group 2009). Несмотря на достижения в геномных технологиях, по-прежнему трудно отобрать множественные вариабельные и ортологичные ядерные маркеры с эквивалентной таксономической широтой и глубиной (но см. Brito and Edwards 2009). Кроме того, для некультивируемых организмов отбор проб с несколькими маркерами у отдельных лиц (в отличие от объединенных проб из окружающей среды в смысле Venter et al. 2004) является сложной задачей (Barraclough et al. 2009). Эволюционный вывод на основе данных одного локуса имеет ограничения, в том числе меньшую мощность для обнаружения независимой эволюции по сравнению с многолокусными подходами (Knowles and Carstens 2007; Dupuis et al. 2012), потенциальное несоответствие между деревьями генов и деревьями видов (Hailer et al. 2012) и отсутствие информации об адаптивной дивергенции (Will and Rubinoff, 2004). Тем не менее, данные по одному локусу действительно обеспечивают генетическую запись эволюционной истории (Avise 2009). ) и, следовательно, предоставляют полезную информацию для изучения эволюционных моделей разнообразия в широком масштабе (Monaghan et al. 2009).
Было предложено несколько методов, подходящих для определения границ видов из данных по одному локусу. Самый простой подход заключается в определении видов на основе правила процентного отсечения, такого как правило 1% или 3%, используемое для разграничения бактериальных видов от последовательностей 16S рДНК (Schloss and Handelsman 2006) или цитохромоксидазы I для видов насекомых (Brower 1994). ). Этот метод страдает от слабой связи с эволюционной теорией, от вариаций типичных уровней внутривидовой и межвидовой изменчивости среди клад и от вариации скорости замещения среди клонов (Barraclough et al. 2009).). Хотя пороги парных расстояний часто могут хорошо работать на практике (Tang et al. 2012), для проверки их использования необходимы эволюционные методы. Кроме того, необходима лежащая в основе эволюционная модель, чтобы установить неопределенность в результирующем разграничении и статистически сравнить альтернативные эволюционные гипотезы, например, диверсифицировалась ли клада в виды или нет (например, Fontaneto et al. 2007).
Эволюционные методы были сосредоточены на обнаружении генетических признаков, указывающих на независимую эволюцию, таких как доказательства фиксированных различий (Hey 1991; Davis and Nixon 1992) или реципрокная монофилия (Wiens and Penkrot 2002) между популяционными выборками. Однако эти подходы требуют априорных гипотез о предполагаемых группировках видов, основанных на традиционной таксономии, морфовидах или выборках популяций, по которым затем можно оценить такие критерии, как монофилия. Это может привести к систематической ошибке в разграничении (например, криптические симпатрические виды не будут разграничены, потому что не будет независимых данных для суждения о монофилии или фиксированных различиях), а необходимая дополнительная информация недоступна в крайних случаях, таких как разграничение бактериальных видов из одного образец окружающей среды исключительно из данных о последовательности ДНК (Acinas et al. 2004; Venter et al. 2004). В более поздних исследованиях были предложены байесовские методы разграничения видов с использованием многолокусной последовательности без априорных границ видов (O’Meara 2010; Yang and Rannala 2010). Эти подходы могут справиться с неопределенностью делимитации и использовать многолокусные данные. Однако в настоящее время они слишком требовательны к вычислительным ресурсам, чтобы их можно было применять к большим выборкам.
Метод Generalized Mixed Yule Coalescent (GMYC), разработанный T.G.B. и впервые представлен в Pons et al. (2006) и Fontaneto et al. (2007), является одним из методов, разработанных для разграничения независимо развивающихся видов с использованием данных по одному локусу. Под «независимой эволюцией» мы подразумеваем, что новые мутации, возникающие у одного вида, не могут легко распространиться на другие виды (Templeton 1989; Barraclough et al. 2003; De Queiroz 2007). Метод GMYC основан на предсказании того, что независимая эволюция приводит к появлению отдельных генетических кластеров, разделенных более длинными внутренними ветвями (Barraclough et al. 2003; Acinas et al. 2004). Он разграничивает такие генетические кластеры, оптимизируя набор узлов, определяющих переходы между межвидовыми и внутривидовыми процессами. Оптимизация продолжается путем нахождения решения максимального правдоподобия (ML) для модели, которая сочетает разнообразие между видами (на основе модели Юла, Nee et al. 19).94) и генеалогическое разветвление внутри вида (на основе нейтрального слияния, Hudson 1990). Поскольку он не опирается на дополнительные данные для формулирования гипотез о предполагаемых видах, этот метод может применяться в случаях отсутствия дополнительных данных, с помощью которых можно было бы сделать вывод о пределах предполагаемых видов. Были предложены и другие методы, основанные на прогнозах, аналогичных тем, которые используются GMYC, и они часто приводят к аналогичным результатам (например, метод K/theta Бирки и др., 2010). Преимущество GMYC заключается в том, что структура правдоподобия позволяет делать статистические выводы и проверять гипотезы по всей отобранной кладе. Относительная эффективность GMYC и альтернативных методов сравнивается в другом месте (Birky et al. 2010; Tang et al. 2012).
Метод был применен к данным одного локуса многих организмов (например, Fontaneto et al. 2007; Lahaye et al. 2008; Papadopoulou et al. 2008), и были предложены расширения алгоритма (Monaghan et al. 2009; Пауэлл 2012). Однако этот метод ранее не был полностью описан или его эффективность не оценивалась по смоделированным данным в широком диапазоне условий (хотя см. Papadopoulou et al. 2008 о влиянии ограниченного рассредоточения и Esselstyn et al. 2012 о влиянии различных эффективный размер популяции и скорость видообразования). Здесь мы подробно представляем используемые алгоритмы и определяем мощность и частоту ошибок для наборов данных, смоделированных в широком диапазоне условий. Кроме того, мы описываем статистические свойства метода разграничения GMYC и вносим коррективы в его формулировку. Пороговое время, используемое для указания местоположения узлов, определяющих виды, теперь правильно обрабатывается как ограничение модели, а не (ошибочно) как параметр. Это изменение не влияет на разграничение ML, найденное с помощью метода, но влияет на способность отвергать нулевую модель, согласно которой все особи принадлежат к одному видовому кластеру.
Методы
Предположения
Предположим, что один локус был секвенирован для выборки особей из монофилетической клады. Образца достаточно, чтобы было отобрано несколько особей для каждого вида, если присутствуют разные виды, и чтобы были отобраны образцы большинства, если не всех видов. Различные схемы выборки будут рассмотрены ниже. Предположим, что истинная генеалогия локуса известна (т. е. отсутствуют артефакты реконструкции), что локус является нейтральным маркером и что внутри каждого вида нет географической подструктуры. Отклонения от этих предположений будут рассмотрены ниже. Цель состоит в том, чтобы определить, диверсифицировалась ли клада на независимо развивающиеся группы, а именно виды, и, если да, разграничить эти виды.
Нулевая модель заключается в том, что все особи в выборке принадлежат к одному виду или популяции. Ожидаемые закономерности для генных деревьев в одной популяции хорошо известны. Выбранные гены сливаются с одним общим предком со средней скоростью, пропорциональной эффективному размеру популяции, N e , в случае строго нейтрального слияния (Hudson 1990; Rosenberg and Nordborg 2002). Слияние происходит потому, что размер популяции ограничен: вероятность того, что каждая особь внесет свой вклад в последующие поколения, зависит от вклада других особей в популяции.
Наша альтернативная модель состоит в том, что клада диверсифицировалась в отдельные виды, каждый из которых рассматривается как единое целое в смысле, изложенном для нулевой модели. Копии гена имеют тенденцию сливаться с одним общим предком внутри каждого вида. Если время до появления самого последнего общего предка ( Tmrca ) внутри вида меньше, чем время, прошедшее с момента отделения вида от его ближайших сестер, это приведет к структуре генетических кластеров: кластеры близкородственных особей, отделенные от других таких кластеров более длинные внутренние ветви (рис. 1; Barraclough et al. 2003). Скорость ветвления между кластерами будет отражать события видообразования, а также вымирание и степень выборки видовых единиц (Nee et al. 19).94; Барраклаф и Херниу, 2003). Ветвление внутри кластеров будет отражать те же популяционные процессы, описанные для нулевой модели, но теперь происходящие независимо у отдельных видов. Однако, если Tmrca внутри вида, как правило, старше, чем время, прошедшее после каждого отделения от ближайшего сестринского вида, то кластеры могут не наблюдаться, и мы не сможем отвергнуть нулевую модель отсутствия разнообразия видов. Обратите внимание, что наше определение сходно с общей концепцией родословной видов, хотя и отличается сосредоточением внимания на генетических сигнатурах, которые мы используем для разграничения таких групп (De Queiroz 2007).
Рисунок 1
Открыть в новой вкладкеСкачать слайд
Схематическая диаграмма, показывающая, как узлы используются для определения видов на гипотетическом дереве генов. Черные ромбы обозначают узлы MRCA, которые определяют виды в конкретном разграничении. Ветви, показанные пунктирной и сплошной линиями, представляют собой ветви видообразования и слияния соответственно. Полосы рядом с наконечниками представляют два альтернативных разграничения. Сплошные полосы обозначают разграничение, определяемое узлами MRCA, показанными на дереве. Черные пунктирные полосы указывают на альтернативное разграничение (т. е. разделение основных видов на два), которое может быть определено многопороговым методом, но не однопороговым. Пунктирные серые вертикальные линии показывают время ожидания ×1, ×2 и т. д., которые используются в модели вероятности GMYC.
Приведенная выше схема предполагает два крайних случая: одна неструктурированная популяция против диверсификации на две или более популяции, каждая из которых неструктурирована, но полностью изолирована от других. На самом деле, у большинства видов также часто присутствует некоторая степень географической структуры (Avise 2009). Это может привести либо к сигнатурам, промежуточным между двумя крайними случаями, либо к иерархической кластеризации, когда кластеры популяций очевидны внутри кластеров видов (Lohse 2009; Papadopoulou et al. 2009).). Имеются выражения для сценариев, отклоняющихся от предположений о нейтральности, постоянном размере популяции и неструктурированной популяции (например, Charlesworth et al. 2003). На данный момент мы игнорируем эту сложность и обратимся к тому, как обнаружить существенные признаки диверсификации в простом случае. Обратите внимание, что этот вопрос не влияет на применение метода для разграничения видов в пределах одной пробы окружающей среды, где не может быть географического подразделения внутри вида.
Вероятностная модель
Подход GMYC расширяет существующие методы правдоподобия для анализа времени событий ветвления в генных деревьях. Необработанными данными для сравнения различных моделей являются времена ожидания между последовательными событиями ветвления, x i (рис. 1). При нулевой модели, в которой вся клада представляет собой единую популяцию (Hudson, 1990), вероятность времени ожидания i в нейтральной коалесцентной модели с эффективным размером популяции N e и n представленных линий определяется как:
Альтернативная модель состоит в том, что клада диверсифицировалась в k видов, каждый из которых рассматривается как независимая популяция с эффективным размером, N j ,j = от 1 до к . Модель разбивает дерево на два типа событий ветвления. Во-первых, события ветвления внутри вида определяются k независимыми нейтральными процессами слияния (показаны черными ветвями на рис. 1). Во-вторых, ветвление между видами (показано пунктирными ветвями на рис. 1), которое мы называем диверсификацией, трактуется как стохастический процесс рождения-гибели, разработанный для анализа видообразования и вымирания на филогенетических деревьях на уровне видов (Nee et al. 1994; Морлон и др. 2011 г.; Этьен и др. 2012). По соглашению, самой простой моделью является модель Юла, в которой число видов растет экспоненциально с течением времени, с постоянным средним коэффициентом рождаемости, λ spec и отсутствием вымирания. Вероятность времени ожидания между последовательными событиями ветвления между видами составляет:
Мы немного модифицировали процесс диверсификации, чтобы можно было сравнить нулевую и альтернативную модели. По соглашению, модели филогенетического ветвления имеют перспективу, начиная с корня и имея M -2 события видообразования, где M — количество типичных точек. Коалесцентные подходы имеют обратную перспективу, начиная с кончиков и имея M -1 коалесцентных событий. Следовательно, если бы мы использовали подход с опережением времени для части диверсификации нашей смешанной модели, у нас было бы на одно событие меньше, чем в нулевой модели, когда выборка берется из одной совокупности. Чтобы учесть это, мы применяем обратный подход к процессу ветвления диверсификации, то есть начинаем со стволовых ветвей для каждого вида, используя уравнение (3) как вероятность того, что произошло предыдущее событие ветвления x i единиц времени ранее (см. Hey 1992). Еще одна сложность заключается в том, что стволовые ветви, ведущие к каждому виду, имеют другое распределение вероятностей, чем ветви диверсификации или слияния, потому что (в обратном времени) каждая стволовая ветвь начинается с общего узла-предка для объекта и заканчивается в событии диверсификации. Самое простое приближение, которое мы принимаем, состоит в том, чтобы предположить, что эти ветви имеют то же распределение вероятностей, что и процесс ветвления диверсификации.
Последним шагом является вычисление вероятности наблюдаемого времени ожидания на дереве, предполагая смешанную модель слияния внутри видов и диверсификации между видами. Комбинированный ряд событий, являющийся результатом комбинации k независимых пуассоновских процессов, соответствует пуассоновскому процессу со скоростью b , равной сумме скоростей отдельных процессов (Кокс и Миллер, 1965, стр. 154). Следовательно, для процесса диверсификации Йоля и набора k нейтральных процесса слияния в рамках заданного распределения видов, вероятность каждого времени ожидания равна:
, где λ spec и n spec используются для обозначения процесса диверсификации, а n i,j — количество линий в интервале ожидания i , принадлежащих процессу j . Расчет соответствующего количества родословных для времени ожидания должен учитывать, какие родословные в любой момент времени принимают участие в каком процессе ветвления (рис. 1). Модель подбирается путем максимизации суммы логарифмических вероятностей времени ожидания для обоих классов событий ветвления по всему дереву. Обратите внимание, что, как и в предыдущих подходах к моделированию процессов диверсификации (Hey 1992), мы не моделируем топологию дерева генов (ср. Rannala and Yang 2003), а вместо этого моделируем распределение интервалов ожидания между событиями ветвления.
Обобщенная модель
Вышеупомянутые уравнения делают строгие предположения о постоянной скорости ветвления как в процессах популяции, так и в процессах диверсификации, которые вряд ли будут широко применяться. Одним из решений может быть сравнение большого количества конкретных моделей населения и диверсификации с использованием информационного критерия Акаике (AIC, Rabosky and Lovette, 2008). Например, можно добавить ветвящийся процесс рождения-смерти для диверсификации, заменив уравнения из Nee et al. (1994). Это упражнение может быть полезно для конкретных гипотез, представляющих предварительный интерес, но оно громоздко из-за огромного количества возможных моделей, многие из которых могут быть неразличимы (Кубо и Иваса, 1995; Барраклаф и Ни, 2001). Альтернативное решение состоит в том, чтобы обобщить приведенные выше уравнения для соответствия ряду качественно различных моделей без указания лежащих в их основе процессов. Ни и др. (1994) и Ни (2001) обсуждали преобразования интервалов ожидания, которые позволяют рассматривать производные модели как экспоненциальные процессы с одним параметром скорости. Эти счета подразумевают простое общее выражение для вероятности времени ожидания одного ветвящегося процесса:
, где p — параметр масштабирования. Значение p = 1 указывает на постоянную скорость ветвления для каждой линии, p > 1 указывает на то, что кажущаяся скорость ветвления увеличивается по направлению к кончикам, а p < 1 указывает на очевидное замедление скорости по направлению к кончикам. Для процессов слияния внутри сущностей n i заменяется на n i ( n i – 1).
Значение p оптимизируется при подборе модели, что позволяет использовать ряд качественно различных моделей. Интерпретация p зависит от того, какой класс событий ветвления рассматривается. Barraclough and Nee (2001) рассмотрели интерпретации изменений скорости диверсификации на филогенетических деревьях, которые точно применимы к интерпретации p . Например, p > 1 указывает на явное увеличение степени диверсификации в сторону настоящего, чего можно было бы ожидать при реальном увеличении рождаемости или в модели с постоянной рождаемостью, но с добавлением постоянной смертности (Nee et al. 1994). p < 1 представляет собой очевидное снижение скорости диверсификации по направлению к настоящему времени, что согласуется с моделями заполнения ниш, или неполной выборкой видов внутри клады (Pybus and Harvey 2000; Nee 2001; Phillimore and Price 2008; Rabosky and Lovette 2008) .
Аналогично, p для модели слияния будет отражать процессы, влияющие на синхронизацию событий слияния внутри сущностей. p < 1 указывает на дефицит недавних событий слияния, например, если популяции увеличиваются в размере или если маркер подвергся выборочному зачистке. p > 1 указывает на избыток недавних событий слияния, например, если популяции сокращались в размерах, если имело место географическое подразделение внутри вида или в результате уравновешивающего отбора по маркеру. (Обратите внимание, что описание в Pons et al. 2006 интерпретации эффектов отбора на слияние p было неверным.) Следовательно, путем оптимизации возможных значений p для обоих классов событий ветвления наш метод качественно устойчив к детали моделей. Обратите внимание, что преобразование мощности может не обеспечивать линейную аппроксимацию конкретного процесса. Тем не менее, он позволяет подобрать плавные изменения скорости ветвления с течением времени, а значения подобранных параметров указывают, какие подробные модели могут быть изучены в качестве возможных объяснений данных.
Используя обозначение, описанное ранее, модель GMYC определяется путем замены b в уравнении (5) на b * :
Оценка ML λ для заданного назначения узлов рассчитывается с использованием Оценка Морана Nee (2001), а именно количество событий ветвления, деленное на общую длину межвидовых ветвей. Оценщик модифицируется, чтобы включить масштабирование длин ветвей, то есть количество событий диверсификации, деленное на сумму ( n i , спец. ) p спец. x i во всех интервалах ожидания. Эквивалентное выражение, но с использованием ( n i,j ( n i,j – 1)) p j x i для деноминатора . Параметры масштабирования оптимизируются с использованием оптимизации Нелдера-Мида, реализованной в функции «optim» R.
Простейшая формулировка состоит в том, чтобы предположить, что все объекты выборки имеют одинаковые параметры ветвления, что мы называем минимальной моделью. В действительности разные включенные в выборку объекты могут иметь разные параметры, если размеры их населения различаются или они имеют разный демографический или селективный опыт. Максимальная модель будет включать отдельные λ j и p j для каждого объекта. Более скромный подход состоит в том, чтобы разрешить варьирование значений параметров между сущностями, но ввести лишь несколько дополнительных параметров в минимальную модель. Мы попытались подобрать отдельные параметры для каждого кластера, и результаты, похоже, не сильно отличаются (см. Pons et al. 2006), хотя в будущих работах этот вопрос может быть изучен подробнее. Для краткости и простоты мы рассматриваем здесь только минимальную версию модели.
Однопороговый подход к разграничению видов
Разграничение с помощью подхода GMYC основано на отнесении событий ветвления к двум категориям, видообразованию и слиянию внутри вида. С упрощающим предположением, что виды являются монофилетическими, можно указать набор узлов самого последнего общего предка (MRCA), который определяет тип событий ветвления (рис. 1). Ветви, происходящие от узлов MRCA, представляют собой сросшиеся ветви внутри вида, а клады, определяемые каждым узлом MRCA, представляют собой видовые кластеры. Поскольку данный набор узлов MRCA определяет форму функции правдоподобия, уравнение (7), и представляет собой уникальную проверяемую гипотезу о принадлежности предполагаемых видов, каждый набор MRCA можно рассматривать как модель-кандидат на определение границ. Модели сравниваются, чтобы получить лучший набор узлов MRCA в схеме ML. Уравнения (6) и (7) сначала оптимизируются отдельно для каждой модели, затем сравниваются значения правдоподобия результатов оптимизации. Набор MRCA с самым высоким ML выбирается как лучшая модель разграничения. Этот процесс аналогичен двухэтапному процессу филогенетического вывода с ML, который рассматривается как выбор модели, а не как оптимизация параметров (Yang et al. 19).95; Ян 2006).
Проблема заключается в том, что даже при упрощающем предположении о монофилии и генетическом дереве небольшого размера существует огромное количество возможных моделей разграничения кандидатов (примерно 4,11×10 22 моделей для сбалансированного дерева со 128 вершинами, онлайн Приложение 1). Самый простой подход, предложенный Pons et al. (2006), состоит в том, чтобы предположить, что существует пороговое время, T , до которого все узлы отражают события диверсификации, а после — все узлы отражают события слияния. Это уменьшает количество моделей-кандидатов до числа узлов в дереве. Он не предполагает, что все виды имеют одинаковые Tmrca , которая будет нарушена даже в модели равного размера популяции из-за ожидаемой дисперсии Tmrca . Вместо этого он предполагает, что самое последнее событие диверсификации произошло до самого старого события слияния внутри вида. Если для экономии предположить, что все виды имеют одинаковую эффективную численность популяции, эта версия модели GMYC вводит два дополнительных параметра по сравнению с нулевой моделью: параметр скорости диверсификации, λ spec и параметр масштабирования p spec . Пороговое время, T , которое рассматривалось как параметр в Pons et al. (2006), теперь корректно рассматривается как ограничение пространства поиска. Порог ML и, следовательно, присвоение вида определяется путем вычисления вероятности альтернативной модели для всех возможных значений T . Сравнение мультимоделей с AIC можно использовать для оценки относительной важности альтернативных моделей и того, предпочтительна ли альтернативная модель ML по сравнению с нулевой моделью отсутствия такого сдвига в процессе ветвления (Powell 2012). Обратите внимание, что с переформулировкой T в качестве ограничения, а не параметра, то степени свободы для сравнения логарифмического правдоподобия нулевой модели с однопороговой моделью теперь равны 2, а не 3, как предложено в Pons et al. (2006). Решение ML не изменилось с новой формулировкой, но мощность такова: любой результат, использующий прежнюю формулировку со значением P 0,112 или меньше, будет значим при 0,05 с пересмотренной моделью.
Многопороговый подход к разграничению видов
Многопороговый подход, разработанный TF и впервые описанный Monaghan et al. (2009 г.), ослабляет предположение однопороговой версии о том, что события видообразования старше, чем все события слияния в генетическом дереве. Вместо этого, исходя из заданного начального назначения узлов MRCA, он ищет альтернативные модели с помощью эвристического алгоритма, который итеративно разделяет и объединяет существующие кластеры видов (см. рис. 1 в Monaghan et al. 2009). Алгоритм продолжает исследовать дополнительные наборы, пока последний раунд испытаний не найдет наборов MRCA с повышенной вероятностью для предыдущего набора ML. Мы попробовали две версии этого процесса, различающиеся тем, как выбирается начальный набор узлов MRCA. Во-первых, мы начали с произвольных наборов, как описано в Monaghan et al. (2009 г.). В этом случае оптимизацию часто привлекают локальные оптимумы, что требует повторений с несколькими начальными наборами, что значительно увеличивает время выполнения. Во-вторых, мы начали с набора MRCA, найденного однопороговым методом. Чтобы проверить эффективность этих подходов, мы смоделировали 100 генных деревьев, предполагая постоянную скорость видообразования и размер популяции 10 90 569 5 90 570 (как описано ниже) с четырьмя видами и пятью верхушками на вид; достаточно мала, чтобы можно было провести исчерпывающий поиск по всем моделям делимитации. Это моделирование показало, что множественный порог, начиная с наилучшего однопорогового решения, чаще находит глобальные оптимумы и лучшие решения, чем поиск, начиная с нескольких произвольных наборов (дополнительный материал в Dryad, doi: 10.5061/dryad.0hv88, таблица S1, рис. С1). Поэтому мы используем этот подход.
В Monaghan et al. (2009), мы рассматривали дополнительные пороги, необходимые для назначения классов в многопороговом алгоритме, как дополнительные параметры, но здесь мы переопределяем их как ограничение пространства поиска, а не параметры, по причинам, описанным ранее. Таким образом, количество параметров теперь одинаково как в однопороговой версии, так и в версии минимальной модели многопороговой версии. Две версии больше нельзя сравнивать с помощью тестов отношения правдоподобия, поскольку они содержат одинаковое количество параметров. Вместо этого альтернативные задания рассматриваются вместе в рамках мультимодельного сравнения.
Неопределенность в разграничении
Мы определяем неопределенность в назначении, используя подход на основе AIC для мультимодельного сравнения, описанный Powell (2012). Веса Акаике и усредненные по модели оценки параметров GMYC и их стандартные ошибки определены в Powell (2012, уравнения 4–9). Кроме того, α % доверительный набор моделей-кандидатов может быть получен путем кумулятивного добавления весов Акаике, начиная с минимальной модели AIC с возрастающим порядком до α 9достигается 0,094 % (Burnham and Anderson 2002). Значение поддержки кластера видов, которое мы называем поддержкой GMYC, определяется суммой весов Акаике моделей, в которых узел MRCA появляется следующим образом.
, где w i — вес Akaike i -й модели, M i и R — общее количество рассматриваемых моделей. Обратите внимание, что это представляет поддержку узла среди альтернативных моделей разграничения, рассматриваемых при сравнении моделей. Хотя не все возможные модели делимитации можно сравнить, сравнение моделей, заданных всеми возможными одиночными порогами, вместе с дополнительными моделями, найденными с помощью многопорогового алгоритма, является разумным подходом.
Моделирование
Для оценки свойств метода генеалогии были смоделированы при нескольких сценариях эволюции и схемах выборки, которые, как ожидается, повлияют на эффективность метода. В каждом последующем варианте, кроме А, деревья видов сначала моделировались в рамках различных моделей диверсификации, а затем моделировались генеалогии генов внутри деревьев видов. Для каждого сценария было смоделировано 100 реплицированных генных деревьев. Деревья видов были смоделированы в Phylogen (Rambaut 2002), а деревья генов — в ms (Hudson 2002), с использованием сценариев в R для создания командных файлов ms, определяющих филогенетическую историю среди видов на основе смоделированного дерева видов. Мы модифицировали ms для вывода длин ветвей до семи знаков после запятой вместо трех, чтобы избежать моделирования одновременных событий слияния для изучаемых здесь больших выборок.
(A) Нулевая модель: Нулевая модель была смоделирована в предположении нейтрального процесса слияния в одной популяции и выборке из 150 особей.
Длина ветвей полученных деревьев выражена в единицах 4 N e поколений. Поскольку метод GMYC опирается на относительную длину ветвей, а не на абсолютную длину ветвей, мы масштабировали длины ветвей в смоделированных деревьях, чтобы иметь возраст корня 1,0 перед запуском анализа GMYC, чтобы поддерживать согласованный масштаб в различных симуляциях.(B) Диверсификация (объединение внутри дерева видов): Было смоделировано генетическое дерево, предполагающее наличие 30 независимо развивающихся видов. Во-первых, видовые деревья были смоделированы в рамках модели постоянного рождения (Йула) без вымирания. Деревья видов были масштабированы таким образом, чтобы возраст корневого узла составлял 10 миллионов поколений, что дало чистую скорость видообразования на линию примерно (log(30)-log(2)) / 10 7 = 0,27, что соответствует типичным значениям. (Барраклаф и Фоглер, 2002). Затем были смоделированы генные деревья внутри видовых деревьев, предполагая, что от каждой сущности было отобрано пять особей.
Числа были выбраны, чтобы соответствовать реалистичным, но консервативным размерам выборки (например, Pons et al. 2006). Влияние выборки большего количества особей на вид по сравнению с выборкой большего количества видов исследуется с помощью моделирования в другом месте (Аренс Д., Краммер Х.Дж., Фудзисава Т., Фабризи С., Фоглер А.П., неопубликованные данные). Численность популяции для каждого вида предполагалась одинаковой и в повторных моделированиях устанавливалась равной 10 9 .0569 4 , 10 5 , 5×10 5 и 10 6 по очереди. Большие размеры популяции увеличивают генетическую изменчивость внутри вида и, следовательно, ожидается, что переход от межвидовых к внутривидовым процессам будет труднее обнаружить. Используемые здесь размеры популяций были выбраны на основе предварительных симуляций, чтобы отражать тенденцию от взаимной монофилии всех видов к случаю, когда несколько видов монофилетичны на генных деревьях с размером популяции 5×10 5 лучше отражает эту тенденцию, чем просто степени 10. Диапазон размера популяции охватывает предполагаемый эффективный размер популяции обычных эукариотических видов (Charlesworth 2009). Обратите внимание, что производительность метода GMYC будет зависеть от относительной скорости диверсификации и слияния, а не от абсолютных скоростей: например, удвоение размера популяции и уменьшение вдвое скорости диверсификации не изменит форму результирующих генных деревьев.(C) Альтернативные модели диверсификации: Моделирование B предполагает постоянную скорость видообразования и отсутствие вымирания. Однако это предположение часто нарушается в реальных наборах данных (Филлимор и Прайс, 2008; Морлон и др., 2011; Этьен и др., 2012). Мы повторили моделирование с альтернативными моделями диверсификации. В C1 были отобраны 30 видов с дерева, содержащего 50 видов, образовавшихся при постоянной скорости видообразования. Эта модель создает недавний дефицит событий видообразования, ситуация, которая может возникнуть из-за неполной выборки видов или фактического замедления чистой скорости диверсификации с течением времени.
Моделирование C2 было смоделировано с постоянной скоростью видообразования и вымирания, при этом скорость вымирания составляла 30% скорости видообразования. Модель с вымиранием производит недавний избыток событий диверсификации, так называемое «притяжение настоящего» (Nee et al. 19).94). Повторение моделирования с коэффициентом исчезновения 80% привело к более экстремальным результатам в том же направлении, что и здесь. Модель GMYC предназначена для соответствия этим различным процессам диверсификации путем оптимизации значений параметров масштабирования, как описано ранее (например, p spec < 1 для C1 и p spec > 1 для C2). Однако ожидается, что разграничение будет проще в C1, чем в моделировании B, потому что время расхождения терминальных видов будет больше, и труднее в C2, потому что терминальные расхождения будут короче. Слившиеся деревья были смоделированы на деревьях видов как C1, так и C2 с той же популяцией и размерами выборки, что и при моделировании B.(D) Альтернативные модели популяций: растущие или сокращающиеся популяции: Моделирование B предполагает, что виды имели постоянные N e с течением времени. Чтобы проверить способность параметров масштабирования соответствовать различным процессам популяции, мы смоделировали две разные модели. В D1 мы смоделировали генные деревья, предполагая недавний экспоненциальный рост внутри вида. Есть много потенциальных версий, которые можно смоделировать, но мы предположили, что на виды повлияло мгновенное узкое место популяции 9.0093 T b поколений назад (произвольно выбрав T b как половину возраста предпоследнего события диверсификации), за которым последовал экспоненциальный рост до наших дней. Скорость роста была назначена для получения 10-кратного увеличения размера популяции, а серьезность узкого места была выбрана таким образом, чтобы гармонический средний эффективный размер популяции за период роста был равен N e , с повторным моделированием.
с Н e равно 10 4 , 10 5 , 5×10 5 и 10 6 для сравнения с моделированием и равно N e . В D2 мы использовали тот же подход, но с мгновенным ростом популяций T b поколений назад с последующим экспоненциальным спадом. Мы прогнозируем, что D1 должен дать p k < 1 и D2 должны давать p k > 1, что отражает относительный дефицит и избыток недавних событий слияния внутри видов соответственно. Мы также прогнозируем, что разграничение должно быть более точным в D1, чем в D2, потому что в D1 будут иметь тенденцию быть относительно более короткие интервалы ветвления рядом с самым последним общим предковым узлом для вида. В действительности маловероятно, что все виды будут демонстрировать одинаковые популяционные тенденции, но моделирование позволяет сравнивать широкие эффекты альтернативных моделей.(E) Диверсификация с популяциями разного размера: Приведенное выше моделирование предполагает, что размеры популяций у видов одинаковы.
Чтобы исследовать влияние различных размеров популяции, мы смоделировали эффективные размеры популяции, взятые из логарифмически нормального распределения со средними значениями, равными 10 4 , 10 5 , 5×10 5 и 10 6 в последовательных симуляциях. Нам неизвестны какие-либо сравнительные данные об эффективных размерах популяций среди видов, но численность видов обычно подчиняется примерно логарифмически нормальному распределению (Грин и Плоткин, 2007). Численность популяций каждого вида пород деревьев, смоделированных в B, была определена из логарифмически нормального распределения со средним значением, равным 10 4 , 10 5 , 5×10 5 и 10 6 в последовательных симуляциях. Предковым видам присваивалась численность популяции вида с меньшим индексом в представлении в формате Ms.(F) Альтернативная схема выборки: случайная выборка по кладам: даже если альтернативная модель верна, случайная выборка может исказить обнаружение истинных кластеров, если нет предварительных знаний об объектах.
Чтобы проверить эффект случайной выборки, мы смоделировали генные деревья с выборкой в общей сложности 150 особей, но выбирая особей случайным образом в зависимости от принадлежности к виду. Моделирование F1 отражало моделирование B с одинаковыми размерами популяций среди видов и, следовательно, с равной вероятностью того, что выбранный индивидуум принадлежит к любому виду. В F2 мы предположили, что размеры популяции распределяются логарифмически нормально, и виды отбираются пропорционально размерам их популяции, то есть виды с удвоенным размером популяции, вероятно, будут отбираться в два раза чаще. Обратите внимание, что в этом моделировании рассматривается вопрос о том, как доля одиночек в выборках влияет на точность разграничения (Lim et al. 2011), поскольку многие виды случайно отбираются только один раз.(G) Географическая структура внутри вида. Для этого сценария мы предположили, что каждый вид разделился на две популяции на полпути вдоль своей конечной ветви дерева видов и что две особи были отобраны из популяции 1 и три особи из популяции 2.
Затем мы присвоили скорость миграции м , так что N e м имеют случайное равномерное распределение от 0 до 1. Предполагалось, что все виды в данном дереве видов имеют одинаковую скорость миграции, но Для повторных видов деревьев была выбрана разная скорость миграции, чтобы изучить, как производительность метода GMYC зависит от скорости миграции. Для m = 0, две популяции полностью изолированы, и метод должен обнаруживать два отдельных кластера внутри каждого вида. Для более высоких м метод должен постепенно переходить от разграничения популяций как кластеров к разграничению видов.(H) Моделирование последовательности и реконструированные генные деревья: Все модели выше предполагают, что истинная генеалогия выбранных генов известна. В действительности, однако, настоящие генные деревья неизвестны и оцениваются по последовательностям ДНК. Неопределенность вывода дерева может снизить эффективность разграничения.
Чтобы оценить эффект вывода дерева, мы провели моделирование последовательности с последующим выводом дерева и разграничением. Последовательности ДНК моделировали вдоль ветвей генных деревьев в модели В с использованием Seq-Gen (Rambaut and Grassly 19).97), используя модель HKY+G и среднюю частоту мутаций 0,02 на миллион поколений (Brower 1994). Длины последовательностей образцов были взяты из равномерного распределения между 200 и 1800, что соответствует диапазону генов, кодирующих митохондриальные белки (Luo et al. 2011). Генные деревья были выведены из смоделированных последовательностей с использованием RAxML (Stamatakis 2006) со 100 псевдорепликатами начальной загрузки, а затем сделаны ультраметрическими с предположением о молекулярных часах с использованием метода Лэнгли-Фитча, реализованного в r8s (Sanderson 2003). Идентичные последовательности (гаплотипы) были сокращены до одной копии перед внедрением из-за известных проблем для GMYC, вызванных идентичными последовательностями (Monaghan et al. 2009).): концевые ветви нулевой длины приводят к вычислению бесконечного слияния λ .
И однопороговый, и многопороговый алгоритмы были запущены на смоделированных генных деревьях. Доверительный набор 95% был построен для каждого прогона моделирования, сначала используя все модели, определенные одними порогами, а затем добавляя модели, найденные с помощью многопорогового поиска. Размер доверительного набора был записан, а частота ложноотрицательных результатов была измерена как частота ошибочного включения нулевой модели в доверительный набор, когда альтернативная модель была истинной. Когда нулевая модель была истинной моделью, скорость исключения нулевой модели из доверительного набора рассчитывалась как частота ложных срабатываний. В качестве меры точности регистрировали среднюю долю видов, правильно разграниченных с точным соответствием. Описанные выше опорные значения рассчитывались для каждого запуска моделирования, и их средние значения использовались для суммирования неопределенности разграничения. Также были записаны оценки ML количества кластеров и параметров масштабирования как для видообразования, так и для коалесцентной части. Для моделирования H результаты GMYC сравнивались с кластерами, полученными с использованием порога расстояния 2% в дереве с соседним соединением смоделированных последовательностей.
Для изучения влияния формы деревьев на делимитации, отдельно от основных различий между моделями, были рассчитаны несколько индексов формы деревьев как для моделируемых видов, так и для генных деревьев. Среднее время ветвления (Mean T spec ) и статистика γ (Pybus and Harvey 2000) видов деревьев были записаны, чтобы обобщить характер событий видообразования. Статистика γ описывает отклонение дерева от модели постоянного видообразования: γ < 0 указывает на явное снижение скорости видообразования, а γ > 0 указывает на явное увеличение скорости видообразования по направлению к кончикам. Обратите внимание, что параметры масштабирования p соответствуют γ , поскольку оба указывают на отклонение от постоянного видообразования (например, когда γ < 0, ожидается p < 1). Индекс Коллесса ( I colless , Agapow and Purvis 2002) был рассчитан для каждого вида деревьев, чтобы проверить влияние дисбаланса деревьев на делимитации. Среднее Tmrca (среднее значение T mrca ) между видами на генных деревьях использовалось в качестве меры изменчивости внутри популяций. Затем был проведен множественный регрессионный анализ с количеством точных совпадений в качестве переменной ответа и индексами дерева генов и эффективным размером популяции в качестве объясняющих переменных. Важность каждого индекса в прогнозировании производительности выражалась в его весе Акаике для моделей, включающих его. Для моделирования G влияние степени структуры популяции на успех делимитации также оценивалось с помощью регрессионного анализа. Вся обработка и анализ данных выполнялись в R (R Development Core Team 2010) с использованием пакета splits (Ezard et al. 2009).) с помощью пользовательских скриптов, а также пакетов APE и apTreeshape (Парадис и др. , 2004; Бортолусси и др., 2006). Последняя версия GMYC в пакете splits с новой формулировкой параметров (версия ≥1.0–15) доступна по адресу http://r-forge.r-project.org/projects/splits и в Дополнительных материалах, депонированных в Dryad. . Обратите внимание, что R-forge поддерживает только версии, совместимые с последней версией R. В ранее доступных версиях GMYC использовалась предыдущая формулировка порогового времени.
Тематические исследования: Tiger Beetles
Однопороговые версии GMYC были применены к данным Pons et al. (2006) и Pons et al. (2011). Решения сравнивались с решениями, полученными с предыдущими версиями. Понс и др. (2006) отобрали 468 особей жуков-тигров рода Rivacindela на 65 участках в Австралии. Группа Rivacindela в значительной степени не была описана, поэтому образцы были отобраны из 108 локальных наборов особей, чтобы представить их морфологическое и географическое разнообразие. Отбирали до пяти особей по морфовидам на участке. Понс и др. (2006) отобрали 161 особь рода Neocicindela по Новой Зеландии из 13 известных таксонов. На каждом участке было отобрано несколько особей вместе с дополнительными представителями описанных видов, чтобы охватить их известный ареал. Генные деревья были выведены из трех областей митохондриальных генов (субъединица 1 цитохромоксидазы, цитохром b и 16S рРНК плюс соседние области) в обоих исследованиях. В обоих случаях мы использовали датированные генные деревья из исходной статьи.
Результаты
Частота ошибок
Частота ложных срабатываний (отказ от нулевой гипотезы при 95%, если это правда) были α = 0,02 и α = 0,07 для одно- и многопорогового методов соответственно. Таким образом, многопороговый метод несколько менее консервативен, чем однопороговый. Частота ложноотрицательных результатов (принятие нулевой гипотезы, когда альтернативная гипотеза верна) была нулевой или близкой к нулю во всех симуляциях с низким N e (10 4 и 10 5 , рис. 2). Таким образом, оба метода работают с одинаково высокой мощностью, когда эффективный размер популяции невелик по сравнению со временем ветвления между видами. Однако частота ложноотрицательных результатов резко возросла для высоких N e (5×10 5 и 10 6 , рис. 2). Многопороговая модель имела большую мощность во всех случаях, например, отбрасывая нулевую модель в 1,1–2,4 раза больше случаев, чем однопороговая при N e = 10 6 .
Рисунок 2
Открыть в новой вкладкеСкачать слайд
Средняя доля ложноотрицательных результатов для каждого типа моделирования и каждого эффективного размера популяции из выборки из 100 смоделированных деревьев для каждого сочетания типа моделирования и размера популяции. Типы моделирования описаны в разделе «Методы»: B — сценарий постоянной скорости видообразования; C1 – сценарий неполной выборки видов; C2 — сценарий постоянной фоновой скорости вымирания; D1 — растущая популяция; D2 – сокращающаяся популяция; E — переменная численность популяций видов; F1 — случайная выборка по кладе, равные размеры популяции; F2 — случайная выборка по кладе, неравные размеры популяции; G – с географической структурой внутри вида; H получен из предполагаемых деревьев из смоделированных данных последовательности.
Частота ошибок также варьировалась в зависимости от процессов генерации, используемых в симуляциях. Как и предсказывалось, частота ложноотрицательных результатов всегда была ниже в C1 (снижение кажущейся скорости диверсификации = более длительное время расхождения между близкородственными видами) и выше в моделировании C2 (увеличение кажущейся скорости диверсификации = более короткое относительное время дивергенции между близкородственными видами), чем в моделировании. B (постоянная норма диверсификации). Вопреки прогнозам, частота ложноотрицательных результатов была ниже в D2 (недавно сокращающаяся популяция), чем в D1 (недавно растущая популяция) или B. При логарифмически нормальном распределении N e (E), частота ложноотрицательных результатов была намного ниже, чем при эквивалентном моделировании с одинаковыми размерами популяций разных видов. Случайная выборка по ветвям, а не по видам, привела к незначительному снижению частоты ложноотрицательных результатов, когда виды имели одинаковые N e (F1 по сравнению с B, рис. 2), но к увеличению частоты ложноотрицательных результатов, когда виды имели разные N . e (F2 против E, рис. 2). Изменение степени миграции между двумя субпопуляциями внутри каждого вида не сильно повлияло на частоту ложноотрицательных результатов (G, рис. 2). Коэффициенты ошибок реконструированных генных деревьев были сравнимы с таковыми для настоящих генных деревьев, за исключением N e = 10 4 (H, рис. 2), в этом случае они были незначительно выше.
Точность разграничения
Средний процент видов, которые были правильно разграничены с точным соответствием, которое мы называем точностью, снизился с более чем 90% при N e = 10 4 до менее 20% при N e = 10 6 (рис. 3). Точность разграничения незначительно снижалась при использовании многопорогового метода во всех случаях (0,6–0,9раз менее точен, чем однократный порог). Среди различных моделей точность была выше в C1 (уменьшение кажущейся скорости диверсификации), чем в C2 (увеличение кажущейся скорости диверсификации), и в D1 (недавно растущие популяции), чем в D2 (недавнее сокращение популяций) на всех N e , как и предсказывалось. . Очевидное несоответствие между результатами по частоте ошибок и точностью относительной эффективности методов в D1 и D2 объясняется тем, что избыток недавних событий слияния в D2 создал искусственные кластеры внутри вида: нулевая модель легче отвергалась, но только из-за неправильное разграничение кластеров. Точно так же, хотя моделирование E (логнормальный N e ) показали гораздо более низкие показатели ложноотрицательных результатов, чем моделирование B, точность не сильно отличалась и действительно для более высоких N e была ниже, чем в моделировании B (рис. 3).
Рисунок 3
Открыть в новой вкладкеСкачать слайд
Средняя точность разграничения, выраженная как доля правильно разграниченных видов, по типам моделирования (от B до H, см. легенду к рис. 2) и по четырем эффективным популяциям размеры. Каждая точка суммирует точность по 100 смоделированным деревьям.
На точность реконструкции не сильно повлияла случайная выборка по кладе, а не по видам (B по сравнению с F1 и F2, рис. 3), и она действительно немного выше при низком уровне N e , хотя относительно высокий доля видов была представлена только одной или двумя особями (моделирование F1: количество образцов на вид колебалось от 1 до 18, при этом 3,3% и 8,7% видов представлены 1 и 2 последовательностями соответственно; моделирование F2 количество образцов на вид варьировались от 1 до 67, при этом 18,2% и 15,3% видов представлены 1 и 2 последовательностями соответственно). Точно так же изменение степени миграции между двумя субпопуляциями внутри каждого вида не сильно повлияло на точность по сравнению с эквивалентным моделированием, предполагающим отсутствие структуры внутри вида (G против B, рис. 3).
В моделировании H многие последовательности пришлось удалить как идентичные гаплотипы из-за низкой изменчивости. Когда N e было 10 4 , 67% последовательностей были удалены, а 49% видов были представлены только одиночными последовательностями (одиночками): это имело самую низкую точность среди всех типов моделирования, но точность все еще была выше 80%. однопороговым методом. Доля идентичных последовательностей и одиночных последовательностей уменьшалась по мере увеличения N e (одиночные последовательности = 5% и идентичные = 33% для N e = 10 5 . Синглтон ≪ 0,01% и идентичный = 7% для N e = 10 6 ), и в этих симуляциях точность не изменилась по сравнению с анализом истинной генеалогии (H против B, рис. 3). Таким образом, вывод генеалогии из последовательностей ДНК не повлиял на точность, за исключением случаев, когда внутривидовая изменчивость была достаточно низкой, чтобы получить множество идентичных последовательностей.
Неопределенность
Размер набора моделей с доверительной вероятностью 95% увеличился на N e увеличились (рис. S2). Тенденции среди различных симуляций отражали тенденции для точности: симуляции, показавшие более высокую точность, сохраняли меньший доверительный набор. Распределение значений поддержки GMYC также изменилось аналогичным образом среди моделей. Например, при моделировании B с однопороговым методом среднее значение поддержки по узлам составило 0,96 для N e = 10 4 , 0,63 для N e = 10 5 , N e = 5× 10 5 и 0,30 для N e = 10 6 . Это показывает, что метод оценки неопределенности адекватно отражает сниженную производительность модели, когда сдвиг в паттернах ветвления внутри вида по сравнению с между видами менее заметен.
Оценка параметра
Расчетное количество кластеров, то есть предполагаемых видов, хорошо совпало с реальным числом видов для N e = 10 4 во всех симуляциях (рис. 4). Оценки снижались, а их диапазон увеличивался с увеличением размера популяции, а различия между типами моделирования соответствовали указанным выше для точности. Расхождение было самым большим в моделировании D2 из-за ложных кластеров внутри видов, обсуждавшихся выше. Многопороговый метод имел тенденцию незначительно завышать количество кластеров по сравнению с однопороговым методом, даже когда N e были небольшими. Различия между типами симуляции отражали те, о которых сообщалось выше для точности, с более высокой точностью, связанной с лучшими оценками количества кластеров. Вывод дерева не повлиял на количество оцененных кластеров даже с учетом количества синглетонов для N e = 10 4 (рис. 4, З).
Рисунок 4
Открыть в новой вкладкеСкачать слайд
Расчетное количество видов на настоящий вид, полученное с помощью а) однопороговой и б) многопороговой версий GMYC. Результаты показаны для каждой комбинации типа моделирования (от B до H) и эффективного размера популяции. Линия внутри каждого прямоугольника показывает медиану, пределы прямоугольника показывают межквартильный диапазон, а усы/точки обозначают экстремальные значения.
Расчетные параметры масштабирования правильно определили различные процессы диверсификации в моделях C1 и C2, когда N e было низким (рис. 5а). В C1 расчетные значения были меньше 1 (медиана = 0,77, V = 377, P ≪ 0,001 для N e = 10 4 тест Уилкоксона), что соответствует недавнему дефициту событий диверсификации. Оценки для C2 были больше 1 (медиана = 1,12, V = 3404, P = 0,003 для N e = 10 4 критерий Уилкоксона), что соответствует недавнему избытку диверсификации. Для высших N e оценки отклонились от ожидаемых закономерностей и уже не отражают закономерности процесса диверсификации (медиана = 1,16, V = 3509, P < 0,001 для С1, медиана = 1,47, V = 3900, P ≪ 0,001 для C2, N e = 10 6 критерий Уилкоксона): кластеры в C2 и N e = 10 6 , рис. 4) означало, что модель больше не оптимизировала относительный избыток недавних событий диверсификации. Оценки скейлинговых параметров для коалесцентного процесса также совпали с смоделированными популяционными процессами, будучи меньше 1 в D1 и больше 1 в D2 (рис. 5б, медиана = 0,53, V = 0, P ≪ 0,001 для D1 и медиана = 1,16, V = 4579, P ≪ 0,01 для D2, N e = 10 4 Тест Уилкоксона). На эти оценки также повлияло увеличение N e , что привело к значениям ниже 1 для всех симуляций при самых высоких значениях N e (медиана = 0,77, V = 0, P ≪ 0,001 для D1). и медиана = 0,82, V = 782, P ≪ 0,001 для D2, N e = 10 6 Критерий Уилкоксона).
Рисунок 5
Открыть в новой вкладкеСкачать слайд
Расчетные параметры масштабирования для а) процесса ветвления диверсификации и б) процесса ветвления слияния для каждой комбинации типа моделирования (от B до H) и эффективного размера популяции с использованием единого -пороговый метод. Горизонтальная пунктирная линия указывает значение 1, которое ожидается в модели по умолчанию, принятой в моделировании B процесса диверсификации с постоянной скоростью и нейтрального процесса слияния без подструктуры и постоянных размеров популяции. Многопороговые результаты представлены на рис. S3 и следуют схожим шаблонам при моделировании.
Структура популяции
Варьирование степени миграции между двумя субпопуляциями внутри каждого вида не оказало существенного влияния на точность разграничения или любые другие показатели эффективности методов при усреднении по всем моделям (G, рис. 2–5). Хотя незначительное снижение точности наблюдалось в промежуточных значениях N e (рис. 6, 10 5 и 5×10 5 ), значительных эффектов миграции и ее взаимодействия с 9 не наблюдалось.0093 N E О Точности ( Z = 1,16, P = 0,24 для N E M и Z = 0,27, P = 0. 7993 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = 0,794 = Z = 0,27, P и . e и N e m , GLM с биномиальной ошибкой).
Рисунок 6
Открыть в новой вкладкеСкачать слайд
Точность разграничения, представленная как доля видов, которые были правильно разграничены, при моделировании с разным уровнем миграции между двумя субпопуляциями ( м ). Отдельные графики показаны для каждого из четырех эффективных размеров популяции, использованных в моделировании.
Реконструированные генные деревья и сравнение с помощью метода, основанного на расстоянии
Длина последовательности не оказала существенного влияния на точность, когда в регрессионном моделировании учитывались все эффективные размеры популяции ( z = 0,75, P = 0,46, GLM с биномиальной ошибкой (рис. S4). Повышение точности с длиной последовательности происходило слабо, только когда N e = 10 4 , и не было значительного взаимодействия между N e и длиной последовательности. Метод GMYC в целом превосходил кластеризацию на основе расстояния с отсечкой 2% (относительная точность метода 2% по сравнению с GMYC: 72% по сравнению с 84%, когда N e = 10 4 , 77% по сравнению с 75 %, когда N e = 10 5 , 26% против 34%, когда N e = 5×10 5 , и 7% против 15%, когда N e = 10 6 ). GLM показал, что общая точность была значительно ниже для метода порога расстояния 2% ( z = -2,22, P = 0,026).
Общие показатели формы дерева и их влияние на точность разграничения
) составил 71 ± 0,58 (мин = 28, макс = 143). Среднее время ветвления (в среднем T spec ) и среднее время коалесценции (Mean T уголь ) варьировались от 0,13 до 0,49 и от 0,0005 до 0,25 со средним значением 0,31±0,002 и 0,06±0,007 соответственно. Эффективный размер популяции ( N e ), статистика γ и среднее время слияния оказали наибольшее влияние на точность (таблица 1). Они были включены во все модели в пределах доверительной вероятности 95% с использованием единого порога (относительная важность = 1,0), и оценки их наклонов значительно отличались от 0: для каждой метрики точность увеличивалась по мере уменьшения ее значения. Среднее T Уголь не был значимым при множественных порогах. Среднее время ветвления имело низкую относительную значимость (0,29), и его оценка не была значимой. Индекс дисбаланса дерева имел наименьшее относительное значение в обоих случаях, как и следовало ожидать, поскольку метод основан на интервалах ветвления междоузлий, а не на топологии.Таблица 1.
Относительная важность (Σ w i ) переменных формы дерева в объяснении точности разграничения при моделировании, а также их усредненные по модели коэффициенты плюс стандартные ошибки (SE)
| Метод . | Одноместный . | Несколько . | ||
|---|---|---|---|---|
| Σ w i | Σ w i | |||
| N e a | 1 | − 2.78× 10 −6 | 1 | − 3,09×10 −6 |
| 3.15×10 −7 | 3.09×10 −7 | |||
| γ b | 1 | − 0.42 | 1 | − 0.23 |
| 5.95×10 −2 | 6.30×10 −2 | |||
| I colless c | 0. 26 | 7.63×10 −4 | 0.21 | 3.36×10 −4 |
| 3.16×10 −3 | 2.50×10 −3 | |||
| Mean T spec d | 0.29 | −1.29×10 −3 | 0.33 | 5.21×10 −3 |
| 1.33×10 −2 | 1.57 ×10 −2 | |||
| Mean T coal e | 1 | − 8.6 | 0.76 | −2.88 |
| ± 2.33 | ±2.37 | |||
| Метод . | Одноместный . | Несколько . | ||
|---|---|---|---|---|
| Σ w i | Σ w i | |||
| N e a | 1 | − 2.78× 10 −6 | 1 | − 3.09×10 −6 |
| 3.15×10 −7 | 3.09×10 −7 | |||
| γ b | 1 | − 0.42 | 1 | − 0.23 |
| 5.95×10 −2 | 6.30×10 −2 | |||
| I colless c | 0. 26 | 7.63×10 −4 | 0.21 | 3.36×10 −4 |
| 3.16×10 −3 | 2.50×10 −3 | |||
| Mean T spec d | 0.29 | −1.29×10 −3 | 0.33 | 5.21×10 −3 |
| 1.33×10 −2 | 1.57 ×10 −2 | |||
| Mean T coal e | 1 | − 8.6 | 0.76 | −2.88 |
| ± 2.33 | ±2.37 | |||
a Средняя эффективная численность населения, жирным шрифтом выделены переменные с коэффициентом, значительно отличающимся от нуля.
b Гамма-статистика Pybus and Harvey (2000).
c Статистика дисбаланса дерева Коллесса.
d Среднее время дивергенции между видами.
e Среднее время слияния видов.
Открыть в новой вкладке
Таблица 1.
Относительная важность (Σ w i ) переменных формы дерева в объяснении точности разграничения при моделировании и их усредненные коэффициенты модели плюс стандартные ошибки (SE)
| Метод . | Одноместный . | Несколько . | ||
|---|---|---|---|---|
| Σ w i | Σ w i | 1288 | ||
| N e a | 1 | − 2.78×10 −6 | 1 | − 3. 09×10 −6 |
| 3.15×10 −7 | 3.09×10 −7 | |||
| γ b | 1 | − 0.42 | 1 | − 0.23 |
| 5.95×10 −2 | 6.30×10 −2 | |||
| I colless c | 0.26 | 7.63×10 −4 | 0.21 | 3.36×10 −4 |
| 3.16×10 −3 | 2.50×10 −3 | |||
| Mean T spec d | 0. 29 | −1.29×10 −3 | 0.33 | 5.21×10 −3 |
| 1.33×10 −2 | 1.57 ×10 −2 | |||
| Mean T coal e | 1 | − 8.6 | 0.76 | −2.88 |
| ± 2.33 | ±2.37 | |||
| Method . | Одноместный . | Несколько . | ||
|---|---|---|---|---|
| Σ w i | Σ w i | 1288 | ||
| N e a | 1 | − 2. 78×10 −6 | 1 | − 3.09×10 −6 |
| 3.15×10 −7 | 3.09×10 −7 | |||
| γ b | 1 | − 0.42 | 1 | − 0.23 |
| 5.95×10 −2 | 6.30×10 −2 | |||
| I colless c | 0.26 | 7.63×10 −4 | 0.21 | 3.36×10 −4 |
| 3.16×10 −3 | 2.50×10 −3 | |||
| Mean T spec d | 0. 29 | −1.29×10 −3 | 0.33 | 5.21×10 −3 |
| 1.33×10 −2 | 1.57 ×10 −2 | |||
| Mean T coal e | 1 | − 8.6 | 0.76 | −2.88 |
| ± 2.33 | ±2.37 | |||
a Mean effective population size, variables with коэффициент, значительно отличающийся от нуля, выделен жирным шрифтом.
b Гамма-статистика Pybus and Harvey (2000).
c Статистика дисбаланса дерева Коллесса.
d Среднее время дивергенции между видами.
e Среднее время слияния видов.
Открыть в новой вкладке
Тигровые жуки
Два набора данных о тигровых жуках различались по степени неопределенности в разграничении. Количество разграниченных видов (количество кластеров и одиночек) колебалось от 40 до 51 (оценка ML 48) с 95% достоверностью для Rivacindela (рис. S5). Обратите внимание, что доверительные интервалы шире, чем ранее сообщавшиеся доверительные интервалы (47–51), основанные на нашем прежнем методе представления решений в пределах двух логарифмических единиц вероятности решения ML. 9Набор с доверительной вероятностью 5% включал 12 из 467 оцененных моделей делимитации, и 27 из 43 узлов, выбранных для разграничения видов в лучшей модели, имели значения поддержки GMYC, равные 1,0 (среднее значение поддержки GMYC для узлов, включенных в набор с доверительной вероятностью 95%, = 0,76). Напротив, метод разграничил от 17 до 55 видов (оценка ML 45) в Neocicindela на уровне достоверности 95% (рис. S6). Опять же, доверительный интервал был шире, чем полученный ранее (32–51, Pons et al. 2011). Набор с доверительной вероятностью 95% включал 31 из 160 сравниваемых моделей, и ни один из узлов не имел значения поддержки GMYC, равного 1,0 (среднее значение поддержки GMYC для узлов, включенных в 95% доверительный набор = 0,33). Причины различий в силе кластеризации в двух кладах обсуждались в другом месте, но могут включать либо более низкую выборку особей в Neocicindela , либо более сильную географическую структуризацию видов Rivacindela на фрагментированных соленых озерах (Pons et al. 2011).
Обсуждение
Метод GMYC для разграничения кластеров из однолокусных генных деревьев широко использовался для разграничения предполагаемых видов из данных типа штрих-кода ДНК, однако его эффективность ранее не проверялась в широких масштабах на смоделированных данных (но см. Papadopoulou et al. 2008 г. Лозе 2009 г.; Эссельстин и др. 2012 для моделирования конкретных функций). Наш анализ показывает, что версии с одним и несколькими порогами достаточно близки к целевой ошибке 0,05, чтобы быть достаточно консервативными при применении к образцам, которые действительно могут происходить из одного неструктурированного кластера видов. Для приложений, требующих проверки этой нулевой гипотезы, мы рекомендуем либо моделировать деревья в рамках нулевой модели для вычисления точного значения P , либо тест хи-квадрат, сравнивающий однопороговую модель GMYC с нулевой моделью для консервативного P -значение.
Наши симуляции также подтверждают предыдущую интуицию о ключевых параметрах, определяющих мощность и точность GMYC. Когда эффективные размеры популяции были низкими по сравнению со временем дивергенции видов, тогда виды были монофилетическими, и метод точно определял кластеры, независимо от подробных предположений, касающихся процессов диверсификации и популяций. По мере увеличения среднего или дисперсии эффективного размера популяции точность разграничения снижалась. В крайнем случае виды больше не были реципрокно монофилетическими и, следовательно, не могли быть точно разграничены методом (предполагающим реципрокную монофилию). Эти результаты совпадают с выводами Esselstyn et al. (2012), которые сравнили оценки видового богатства для деревьев, смоделированных с различными отношениями N e к скорости видообразования.
Точность варьировалась в зависимости от различных симуляций в основном в соответствии с предсказанными шаблонами, основанными на эффектах конкретных процессов. Параметры масштабирования действительно точно отражали процессы диверсификации и слияния в моделировании, когда размеры популяции были небольшими. Тем не менее, включение более реалистичных макроэволюционных моделей может повысить производительность с реальными данными. Например, эмпирические исследования показали, что фоновая скорость угасания может составлять около 90% скорости видообразования (Ricklefs 2007; Etienne et al. 2012). Это приведет к размытию резкости перехода от межвидового к внутривидовому ветвлению и, следовательно, к снижению точности метода GMYC. Хотя параметр масштабирования мог соответствовать этой тенденции в кажущейся скорости диверсификации, более точное разграничение можно было бы получить, явно включив вымирание в модель. Последнее предположение, касающееся диверсификационной части модели, состоит в том, что темпы диверсификации одинаковы для всех клад. Опять же, модели, допускающие такое отклонение, могут быть реализованы (Morlon et al. 2011).
Производительность метода также зависит от отбора проб несколькими способами. Увеличение выборки особей по видам повысит производительность. Здесь мы смоделировали выборку только из пяти особей на вид, чтобы отразить, вероятно, ограниченную выборку внутри вида для широких исследований биоразнообразия. Увеличение этого числа может увеличить кажущуюся скорость ветвления внутри кластеров и, следовательно, способность метода обнаруживать порог скорости ветвления (см. Reid and Carstens 2012). Например, в исследованиях жуков-тигров среднее число особей на разграниченный вид составляло 9 особей.0,75 (min = 1, max = 63, 10% и 13% кластеров, представленных 1 и 2 последовательностями соответственно) для Rivacindela , в которых были получены высокие значения поддержки, но только 3,58 (min = 1, max = 21, 51% и 9% кластеров представлены 1 и 2 последовательностями соответственно) для Neocicindela , у которых поддержка GMYC была намного ниже. И наоборот, увеличение выборки видов в пределах клады (но при сохранении постоянного числа особей, отобранных для каждого вида) может уменьшить возможность разграничения видов, включенных в выборку. Это происходит потому, что случайное упущение видов снижает вероятность того, что будут отобраны наиболее близкие родственные виды каждого вида, и, следовательно, снижает вероятность обнаружения немонофилетических видов. Этот результат был продемонстрирован эмпирически для увеличения пространственного масштаба выборки европейских водяных жуков (Bergsten et al. 2012). Влияние количества особей на вид по сравнению с количеством отобранных видов на точность метода исследуется с помощью моделирования в другом месте: GMYC может точно работать всего с 3 видами, но он более надежен при более чем 10 видах (Ahrens D. ., Krammer H.J., Fujisawa T., Fabrizi S., Vogler A.P., неопубликованные данные).
Наконец, хотя можно было ожидать, что случайная выборка по кладе по сравнению со случайной выборкой внутри каждого вида повлияет на метод, наши результаты показали, что эффект был небольшим по сравнению с эффектами N e (сравните B и F2 на рис. 3). Доля видов, представленных только одной или двумя последовательностями в этих моделях (12% в F1 и 33,5% в F2), отражает реалистичные уровни редкости во многих исследованиях биоразнообразия (Lim et al. 2011). Этот результат означает, что на метод, вероятно, мало повлияет то, взяты ли образцы равномерно на основе предварительных выводов об ограничениях видов (как во многих исследованиях штрих-кодирования ДНК многоклеточных эукариот) или без такого знания (как в исследованиях некультивируемых бактерий или эукариот из пробы окружающей среды). Наш код моделирования доступен в дополнительных материалах для пользователей, желающих исследовать возможные эффекты их собственного плана выборки.
Мы обнаружили, что ослабление ограничения одного порога не приводит к повышению производительности метода. Многопороговая версия дает довольно схожие результаты с однопороговой версией, но с тенденцией к чрезмерному разбиению. В этом случае узел, восстановленный однопороговой версией, обычно по-прежнему включается в набор узлов MRCA с доверительной вероятностью 95 %, но с меньшей достоверностью. Хотя возможно, что в будущей работе можно будет описать многопороговый алгоритм, который мог бы улучшить разграничение, например, более исчерпывающий поиск с использованием байесовской цепи Маркова Монте-Карло (Рейд и Карстенс, 2012), вместо этого мы полагаем, что это может отражать фундаментальное ограничение потенциал метода. Представьте себе крайний случай, когда у каждого вида было разное время до MRCA и скорость ветвления внутри кластеров. В этом случае не будет среднего сигнала по дереву, по которому можно восстановить сдвиг в процессе ветвления. Только когда сдвиги совпадают на определенном уровне дерева, мы можем найти статистические доказательства кластеризации. Наши результаты не исключают, что версия с несколькими порогами может давать улучшенные результаты в некоторых случаях (что будет означать исключение узла MRCA, обнаруженного в версии с одним порогом). Поэтому мы рекомендуем с осторожностью использовать многопороговую версию как способ изучения того, как меняется разграничение при ослаблении допущений единого порога.
Модель GMYC ранее подвергалась критике за предположение, что виды составляют единую неструктурированную популяцию. Географическая структура является общей чертой многих реальных видов, и в сочетании с неслучайной выборкой ареалов видов это может привести к систематической ошибке при разграничении видов. Предыдущие симуляции для исследования этой проблемы касались способности GMYC разграничивать кластеры вдоль континуума от одной популяции до популяции, структурированной в субпопуляции без потока генов (например, Papadopoulou et al. 2008, 2009).; Лозе 2009). В таких случаях было показано, что GMYC может разграничить частично изолированные популяции как отдельные виды, особенно если не были отобраны связывающие популяции. Однако для большинства реальных применений GMYC более уместно рассмотреть, как географическая структура внутри каждого вида влияет на способность GMYC разграничивать виды в более широкой кладе, включающей несколько отдельных видов. Наше моделирование здесь показывает, что, пока соблюдаются условия для точного разграничения видов (т. низкая вариация внутри каждого вида, порог был оптимизирован на правильном уровне дерева.
Метод GMYC требует предварительного удаления идентичных последовательностей, поскольку концевые ветви нулевой длины затрудняют оценку правдоподобия; то есть оценка Морана λ не может быть правильно оценена из-за нулевой общей длины ветвей в кластерах. Это может быть особенно проблематично, когда генетические деревья субъектов реконструируются из медленно развивающихся маркеров, что приводит к уменьшению общего размера выборки и изменчивости внутри вида. Однако имитационное исследование показало, что метод толерантен к умеренному количеству идентичных последовательностей и одиночек, разграничивая их, корректно приписывая им видообразование. Точность реконструированных генных деревьев, смоделированных для степени изменчивости и диапазона длин маркеров, типичных для мтДНК и других исследований с одним локусом, также была сравнима с результатами, полученными с использованием истинной генеалогии. Кроме того, GMYC в целом работал лучше, чем кластеризация с пороговым значением 2%, что указывает на преимущество варьирования границ видов по сравнению с фиксированным пороговым значением, хотя точность была эквивалентной, когда N e = 10 5 . Относительная эффективность этих методов будет зависеть от типичных уровней N e и различий в коэффициентах замещения: в недавнем исследовании таксонов животных GMYC и использование 3% порога дали аналогичные результаты (Tang et al. 2012). .
Одним из явных ограничений GMYC является то, что он предполагает, что виды являются монофилетическими. Это предположение нарушается для реальных видов, когда времени дивергенции было недостаточно для того, чтобы сущности приобрели монофилию (Hudson and Coyne 2002). Снижение совпадений между истинными сущностями и оценочными кластерами отражает это нарушение. Согласно теоретическим исследованиям время возникновения реципрокной монофилии нейтрального маркера с 95% вероятность между парой видов составляет 8,7 Ne для одного ядерного маркера и 2,2 Ne для мтДНК (Hudson and Coyne 2002). Диапазон немонофилии видов в наших симуляционных исследованиях составлял от 0% до 87% (медиана 20%), что хорошо перекрывало показатель в 23%, полученный в результате метаанализа популяционно-генетических исследований (Фанк и Омланд, 2003). Доля немонофилетических видов сильно коррелировала с точностью разграничения (рис. S7). Прогнозируемая точность для 20% немонофилии составила около 50% точных совпадений.
Признаки недавних событий видообразования, таким образом, не могут быть обнаружены с помощью этого метода (или любого другого метода, использующего монофилию отдельных генов), хотя точность 50 % все же может быть полезной для быстрой оценки видового разнообразия. Это ограничение может быть устранено с помощью многолокусного анализа, который позволяет разграничить недавно дивергировавшиеся сестринские виды (Knowles and Carstens 2007). Однако в настоящее время методы требуют предварительных гипотез о популяциях видов и требуют значительных вычислительных ресурсов для больших размеров выборки. Ключевой задачей многолокусных подходов является поиск эффективных алгоритмов поиска альтернативных границ (Янг и Раннала, 2010). Обратите внимание, что GMYC можно использовать для разграничения кластеров из любых ультраметрических деревьев, в том числе полученных из объединенных многолокусных данных: хотя он будет менее мощным, чем многолокусные методы для половых организмов (Knowles and Carstens 2007), возможно, его можно использовать для поиска начальных разграничений. для оценки с помощью мультилокусных подходов.
Выводы
Метод GMYC изначально был разработан как метод отграничения видов от генных деревьев одного локуса в отсутствие какой-либо дополнительной информации. Это достигается за счет обнаружения генетической кластеризации за пределами уровней, ожидаемых в нулевой модели, согласно которой все отобранные люди принадлежат к одной взаимодействующей популяции. В группах, в которых эффективный размер популяции, как правило, невелик, а время расхождения между видами, как правило, велико, этот метод является точным и консервативным. У многих эукариот и прокариот в деревьях генов для клад или образцов окружающей среды проявляются четкие паттерны генетической кластеризации, что указывает на то, что условия для GMYC часто выполняются. Везде, где доступна дополнительная информация, такая как местонахождение выборки особей, количественная или качественная информация о фенотипических признаках или дополнительные генные последовательности, эта информация должна использоваться для перекрестной проверки объектов, ограниченных GMYC. У более крупных эукариот такая информация обычно доступна, и полезность GMYC заключается в предоставлении объективных средств разграничения генетических кластеров для сравнения с другими данными. Следует приложить усилия для отбора проб особей на репрезентативном ареале, занимаемом видом, и учитывать потенциальные пределы точности метода, описанного выше. Задача, для которой GMYC лучше всего подходит, — это разграничение единиц разнообразия, когда доступны только данные о последовательности ДНК, например, в исследованиях ДНК окружающей среды. Там он обеспечивает эволюционную основу для дополнения существующих прагматических методов, основанных на пороговых значениях, основанных на эмпирическом опыте.
Дополнительный материал
Файлы данных и/или другая дополнительная информация, относящаяся к этому документу, были депонированы в Dryad под номером: 10.5061/dryad.0hv88.
Финансирование
Время TGB было поддержано грантом Совета по исследованиям в области биотехнологии и биологических наук (BBSRC) [BB/G004250/1].
Благодарности
Мы благодарим Майкла Монагана, Йоханнеса Бергстена, Шона Ни и Альфрида Фоглера за обсуждение идей, содержащихся в этой рукописи, Зихенга Янга за обсуждения, приведшие к новой интерпретации порогового параметра, Кевина Балби за выполнение некоторых симуляций, Тома Эзарда. за подготовку разделения пакетов R и Ричарда Глора и четырех анонимных рецензентов за комментарии.
Каталожные номера
Acinas
S.G.
,
Klepac-Ceraj
V.
,
Hunt
3
,
Фарино
C.
,
Ceraj
I.
,
Distel
D.900
,
Polz
М.Ф.
.
Мелкомасштабная филогенетическая архитектура сложного бактериального сообщества
,
Природа
,
2004
, том.
430
(стр.
551
—
554
)
Агапов
П.-М.
,
Пурвис
А.
.
Мощность статистики формы восьми деревьев для обнаружения неслучайной диверсификации: сравнение путем моделирования двух моделей кладогенеза
,
Сист. биол.
,
2002
, том.
51
(стр.
866
—
872
)
Avise
2J.C.
Филогеография: ретроспектива и перспективы
,
Ж. Биогеог.
,
2009
, том.
36
(стр.
3
—
15
)
Barraclough
Т.Г.
,
Бирки
C.W.
Младший
,
Берт
А.
.
Диверсификация половых и бесполых организмов
57
(стр.
2166
—
2172
)
Barraclough
Т.Г.
,
Эрниу
E.
.
Почему существуют виды? Взгляды сексуалов и асексуалов
,
Зоология
,
2003
, том.
106
(стр.
275
—
282
)
Barraclough
Т.Г.
,
Hughes
M.
,
Ashford-Hodges
N.
,
Fujisawa
T.
.
Вывод эволюционно значимых единиц бактериального разнообразия из обширных экологических исследований данных по одному локусу
,
Биол. лат.
,
2009
, том.
5
(стр.
425
—
428
)
Barraclough
Т.Г.
,
.
Филогенетика и видообразование
,
Trends Ecol. Эвол.
,
2001
, том.
16
(стр.
391
—
399
)
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Недавние темпы диверсификации североамериканских тигровых жуков, оцененные на основе датированного филогенетического дерева мтДНК
,
Mol. биол. Эвол.
,
2002
, том.
19
(стр.
1706
—
1716
)
Бергстен
Дж. 0003
Bilton
D.T.
,
Fujisawa
T.
,
Elliot
M.
,
Monaghan
M.T.
,
Balke
M.
,
Hendrich
L.
,
Geijer
J.
,
Herrmann
J.
,
Foster
G.N.
,
Рибера
I.
,
Нильссон
А.Н.
,
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Влияние географического масштаба выборки на ДНК-штрихкодирование
,
Сист. биол.
,
2012
, том.
61
(стр.
851
—
869
)
Birky
C.W.
Jr.
,
Adams
J.
,
Gemmel
M.
,
Perry
J.
.
Использование популяционно-генетической теории и последовательностей ДНК для обнаружения и идентификации видов у бесполых организмов
5
стр.
e10609
Бортолусси
N.
,
Durand
E.
,
Blum
M.
,
Francois
O.
.
apTreeshape: статистический анализ формы филогенетического дерева
22
(стр.
363
—
364
)
Brito
стр.
0222 ,Эдвардс
С.В.
.
Многолокусная филогеография и филогенетика с использованием маркеров на основе последовательностей
135
(стр.
439
—
455
)
Брауэр
А.В.З.
.
Быстрое морфологическое излучение и конвергенция рас бабочек Heliconiuserato выведено из закономерностей эволюции митохондриальной ДНК
,
Proc. Натл. акад. науч. США
,
1994
, том.
91
(стр.
6491
—
6495
)
Бернем
К.П.
,
Андерсон
Д.Р.
. ,
Выбор модели и мультимодельный вывод: практический информационно-теоретический подход
,
2002
New York
Springer Science+Business Media, Inc
Camargo
A.
,
Avila
L.J.
,
Morando
M.
,
Sites
Дж. В. младший
.
Точность и прецизионность деревьев видов: влияние выборки локусов, отдельных особей и пар оснований на вывод о деревьях видов у ящериц группы Liolaemus darwinii (Squamata, Liolaemidae)
,
Сист. биол.
,
2012
, том.
61
(стр.
272
—
288
)
Carstens
B.C.
,
Дьюи
Т.А.
.
Разграничение видов с использованием комбинированного коалесцентного и теоретико-информационного подхода: пример из Северной Америки Myotis летучие мыши
,
Syst. биол.
,
2010
, том.
59
(стр.
400
—
414
)
Рабочая группа завода CBOL
ДНК -шар -код для наземных растений Proc
,
NATL. акад. науч. США
,
2009
, том.
106
(стр.
12794
—
12797
)
Чарльзворт
3
Эффективный размер популяции и закономерности молекулярной эволюции и изменчивости
,
Nat. Преподобный Жене.
,
2009
, том.
10
(pg.
195
—
205
)
Charlesworth
B.
,
Charlesworth
D.
,
Barton
N.H.
.
Влияние генетической и географической структуры на нейтральную изменчивость
,
Анн. Преподобный Экол. Эвол. Сист.
,
2003
, том.
34
(стр.
99
—
125
)
Cox
Д.Р.
,
Миллер
Х.Д.
. ,
Теория случайных процессов
,
1965
London
Chapman & Hall
Coyne
J.A.
,
Орр
Х.А.
.
Эволюционная генетика видообразования
,
Фил. Транс. Р. Соц. Лонд. Б
,
1998
, том.
353
(стр.
287
—
305
)
Койн
Дж.А.
,
Орр
Х.А.
. ,
Спецификация
,
2004
Сандерленд (Массачусетс)
Синауэр Ассошиэйтс. Inc
Davis
J.I.
,
Никсон
К.С.
.
Популяции, генетическая изменчивость и разграничение филогенетических видов
,
Сист. биол.
,
1992
, том.
41
(стр.
421
—
435
)
Де Кейрос
К.
К.
Понятия видов и разграничение видов
,
Сист. биол.
,
2007
, том.
56
(стр.
879
—
886
)
DUPUIS
J.R.
,
.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H.H…
222,
222,
222,
222,
9.h.h.h.h.h.h.h.h.h.h.h.h.h.h.h.h.h.h.
,
..
Разграничение многолокусных видов у близкородственных животных и грибов: одного маркера недостаточно
,
Mol. Экол.
,
2012
, том.
21
(стр.
4422
—
4436
)
Эссельстин
Дж.А.
,
Evans
B.J.
,
Sedlock
J.L.
,
Anwarali Khan
F.A.
,
.
.
Разграничение видов по одному локусу: испытание смешанной йольско-слившейся модели с эмпирическим применением к филиппинским круглолистным летучим мышам
,
Проц. Р. Соц. Лонд. Б.
,
2012
, том.
279
(стр.
3678
—
3686
)
Этьен
Р.С.
,
Haegeman
B.
,
Stadler
T.
,
,
Pearson
P.N.
,
Первис
А.
,
Филлимор
А.Б.
.
Зависимость от разнообразия приближает молекулярные филогении к согласию с летописью окаменелостей
,
Proc. Р. Соц. Лонд. Б
,
2012
, том.
279
(стр.
1300
—
1309
)
EZARD
T.
,
Fujisawa
T.
,
,
,
,
,
,
,
,
,
,
,
,
,
,
,
,
9222,.0003
Т.Г.
.
РАЗДЕЛЕНИЕ: Ограничения видов по пороговой статистике. Версия пакета R 1.0-18/r45
,
2009
Feulner
P.G.D.
,
Kirschbaum
F.
,
Mamonekene
V.
,
Ketmaier
V.
,
Tiedemann
.Адаптивная радиация у африканских слабоэлектрических рыб (Teleostei: Mormyridae: Campylomormyrus ): комбинированный молекулярный и морфологический подход
,
J. Evol. биол.
,
2007
, том.
20
(стр.
403
—
414
)
Fontaneto
D.
,
Herniou
E.A.A.
,
Boschetti
C.
,
Caprioli
M.
, 9Т.Г.
.
Независимо развивающиеся виды бесполых бделлоидных коловраток
,
PLoS Biol.
,
2007
, том.
5
стр.
e87
Фанк
Д.Дж.
,
Омланд
К.Е.
.
Парафилия и полифилия на уровне вида: частота, причины и последствия на основе анализа митохондриальной ДНК животных
,
Ann. Преподобный Экол. Эвол. Сист.
,
2003
, том.
34
(стр.
397
—
423
)
Грин
J.L.
,
Plotkin
J.B.B.B.
2,
.
Статистическая теория выборки численности видов
,
Экол. лат.
,
2007
, том.
10
(стр.
1037
—
1045
)
Hailer
F.
,
Kutschera
V.E.
,
Холлстрем
Б.М.
,
Классерт
Д.
,
Файн
С.Р.
,
Леонард
Дж.А.
,
Арнасон
У.
,
Янке
А.
.
Ядерные геномные последовательности показывают, что белые медведи являются древней и самобытной медвежьей родословной
336
(стр.
344
—
347
)
Hebert
P.D.N.
,
Цивинска
A.
,
Мяч
S.L.
,
DeWaard
J.R.
.
Биологическая идентификация с помощью штрих-кодов ДНК
,
Proc. Р. Соц. Лонд. Б
,
2003
, том.
270
(стр.
313
—
321
)
.
Структура генеалогий и распределение фиксированных различий между образцами последовательностей ДНК из природных популяций
,
Генетика
,
1991
, том.
128
(стр.
831
—
840
)
.
Использование филогенетических деревьев для изучения видообразования и вымирания
,
Эволюция
,
1992
, vol.
46
(стр.
627
—
640
)
.
Разум видов проблема
,
Trends Ecol. Эвол.
,
2001
, том.
16
(стр.
326
—
329
)
Хадсон
2 Р. Р.
Футуйма
Д.Дж.
,
Антонивикс
Дж.
.
Генеалогия генов и процесс слияния
,
Oxford Surveys in Evolutionary Biology
,
1990
Oxford
Oxford University Press
(стр.
1
—
44
)
Hudson
R.R.
2222222222222222222222.
Генерация образцов по нейтральной модели генетической изменчивости Райта-Фишера
18
(стр.
337
—
338
)
Hudson
R.R.
,
Coyne
J.A.
.
Математические следствия концепции генеалогического вида
56
(стр.
1557
—
1565
)
Ноулз
L.L.
,
Carstens
B.C.
.
Разграничение видов без монофилетических генных деревьев
,
Сист. биол.
,
2007
, том.
56
(стр.
887
—
895
)
KOUFOPANOU
V.
,
22,
2,
2,
9000. 9000.22,
9000. 9000.22,
9000.22,
9000.2,
9000.2222,9000.
2,
9000.
2.
.
Конкордантность генеалогий генов указывает на репродуктивную изоляцию патогенного грибка Coccidioides immitis
,
Proc. Натл. акад. науч. США
,
1997
, том.
94
(стр.
5478
—
5482
)
KUBO
T.
,
IWASA
y..
222222222222222222.
Определение скоростей ветвления и вымирания на основе молекулярных филогений
49
(pg.
694
—
704
)
Lahaye
R.
,
van Der Bank
M.
,
Bogarin
D.
,
Warner
J.
,
Pupulin
F.
,
Gigot
G.
,
Maurin
O.
,
Duthoit
S.
,
Барраклаф
Т.Г.
,
Саволайнен
В.
.
ДНК-штрихкодирование флор горячих точек биоразнообразия
,
Proc. Натл. акад. науч. США
,
2008
, том.
105
(стр.
2923
—
2928
)
Leaché
A.D.
,
Koo
M.S.
,
Спенсер
К.Л.
,
Папенфус
Т.Дж.
,
Фишер
Р.Н.
,
McGuire
J.A.
.
Количественная экологическая, морфологическая и генетическая изменчивость для разграничения видов в комплексе видов прибрежных рогатых ящериц ( Phrynosoma )
,
Proc. Натл. акад. науч. США
,
2009
, том.
106
(стр.
12418
—
12423
)
LIM
G.S.
,
2.
.
292..
9292
.
9292
.
2.92
.
92
9292.
92
Определение границ видов в мире, полном редкости: снглетоны, методы разграничения видов
,
Сист. биол.
,
2011
, том.
61
(стр.
165
—
169
)
Лозе
К.
.
Можно ли использовать штрих-коды мтДНК для разграничения видов? Ответ Pons et al. (2006)
,
Сист. биол.
,
2009
, том.
58
(стр.
439
—
442
)
,
Zhang
A.
,
HO
с..
,
,
Чжан
Y
,
,
Кэмерон
С.Л.
,
.
Потенциальная эффективность митохондриальных генов для штрих-кодирования ДНК животных: тематическое исследование с использованием плацентарных млекопитающих
12
стр.
84
Мейер
К.П.
,
Поле
Г.
.
Штрих-кодирование ДНК: частота ошибок на основе всесторонней выборки
,
PLoS Biol.
,
2005
, том.
3
стр.
e422
Монаган
М.Т.
,
Дикий
Р.
,
Эллиот
М.
,
Фудзисава 9
003
Balke
M.
,
Внутрь
D.J.G.
,
Lees
D.C.
,
Ranaivosolo
R.
,
Eggleton
P.
,
Barraclough
T.G.G.G.G.G.G.G.
,
Фоглер
А. П.
.
Ускоренная инвентаризация видов на Мадагаскаре с использованием основанных на коалесцентах моделей определения границ видов
,
Сист. биол.
,
2009
, том.
58
(стр.
298
—
311
)
Morlon
H.
,
Parsons
T.L.
,
Плоткин
Дж.Б.
.
Согласование молекулярных филогений с летописью окаменелостей
,
Proc. Натл. акад. науч. США
,
2011
, том.
108
(стр.
16327
—
16332
)
.
Определение скорости видообразования на основе филогений
55
(стр.
661
—
668
)
,
Май
3 Р.
3 ,
Харви
П.Х.
.
Реконструированный эволюционный процесс
,
Фил. Транс. Р. Соц. Лонд. Б
,
1994
, том.
344
(стр.
305
—
311
)
О’Мира
B.C.
.
Новые эвристические методы для совместного разграничения видов и вывода дерева видов
,
Сист. биол.
,
2010
, том.
59
(pg.
59
—
73
)
Papadopoulou
A.
,
Bergsten
J.
,
Fujisawa
T.
,
Монаган
М.Т.
,
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Видообразование и ДНК-штрихкоды: проверка влияния дисперсии на формирование дискретных кластеров последовательностей
,
Фил. Транс. Р. Соц. Лонд. Б
,
2008
, том.
363
(стр.
2987
—
2996
)
Papadopoulou
A.
,
Monaghan
M.T.T.T.T.
,
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Ошибка выборки не делает недействительной модель слияния Юла для разграничения видов. Ответ Лозе (2009 г.)
,
Сист. биол.
,
2009
, том.
58
(стр.
442
—
444
)
Paradis
E.
29222,92
22,92
2,92
2,92
2,92
2. APE: анализ филогенетики и эволюции на языке R
,
Биоинформатика
,
2004
, том.
20
(стр.
289
—
290
)
Phillimore
A.B.
,
Цена
Т.Д.
.
Зависимый от плотности кладогенез у птиц
,
PLoS Biol.
,
2008
, том.
6
стр.
e71
Pons
J.
,
Барраклаф
Т.Г.
,
Гомес-Зурита
Дж.
,
Кардосо
А.
,
Дюран
3
,
Hazell
S.
,
Kamoun
S.
,
Sumlin
W.D.
,
Vogler
A.P.
,
A.P.
,
A. P.
,
A.D.
Разграничение видов на основе последовательностей для таксономии ДНК неописанных насекомых
,
Сист. биол.
,
2006
, том.
55
(pg.
595
—
609
)
Pons
J.
,
Fujisawa
T.
,
Claridge
E.M.
,
Сэвилл
Р.А.
,
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Глубокое подразделение мтДНК у линнеевских видов в эндемической радиации тигровых жуков из Новой Зеландии (род Neocicindela) Фил. Эвол.
,
2011
, том.
59
(стр.
251
—
262
)
Пауэлл
2 Дж. Р. 9003.
Учет неопределенности в разграничении видов при анализе данных о последовательностях ДНК из окружающей среды
,
Методы Экол. Эвол.
,
2012
, том.
3
(стр.
1
—
11
)
Pybus
O.G.
,
Харви
П.Х.
.
Тестирование макроэволюционных моделей с использованием неполных молекулярных филогений
,
Proc. Р. Соц. Лонд. Б
,
2000
, том.
267
(стр.
2267
—
2272
)
R Development Core Team
,
R: A Язык и окружающая среда для статистических вычислений
,
20102 2010 2010 20102 2010 20102 2010 2010 20102 2010 2010 20102 20102 2010 2010 20102 2010 20102 20102 2010 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 20102 2010 2010 2 20102 20102 2010 2010 2 20102 2010 2010 2 20102 2010 2010 2010 2 2010 2 20102 2010 2010 2010 2 20102. )
R Фонд статистических вычислений
Рабоски
Д.Л.
,
Ловетт
И.Дж.
.
Зависимая от плотности диверсификация североамериканских лесных камышевок
,
Проц. Р. Соц. Лонд. Б
,
2008
, том.
275
(стр.
2363
—
2371
)
Рамбо
А.
Phylo-O-Gen: Phylogenetic Tree Simulator Package v1.1
,
2002
Rambaut
A.
,
Gravly
N.C.
222222222222222.
Seq-Gen: приложение для моделирования эволюции последовательностей ДНК вдоль филогенетических деревьев методом Монте-Карло
,
CABIOS
,
1997
, том.
13
(стр.
235
—
238
)
Раннала
B.
,
Yang
Z.
2.
Байесовская оценка времени дивергенции видов и размера предковой популяции с использованием последовательностей ДНК из нескольких локусов
,
Генетика0222 , том.
164
(стр.
1645
—
1656
)
Reid
N.M.
,
Carstens
г. до н.э.
.
Ошибка филогенетической оценки может снизить точность разграничения видов: байесовская реализация модели General Mixed Yule Coalescent
,
BMC Evol. биол.
,
2012
, том.
12
стр.
196
Риклефс
Р.Э.
.
Оценка темпов диверсификации на основе филогенетической информации
,
Trends Ecol. Эвол.
,
2007
, том.
22
(стр.
601
—
610
)
Rosenberg
N.A.
,
Nordborg
M. 9000
2222.
Генеалогические деревья, теория слияния и анализ генетических полиморфизмов
,
Нац. Преподобный Жене.
,
2002
, том.
3
(стр.
380
—
390
)
Sanderson 2
M.J.
3.
r8s: вывод абсолютных скоростей молекулярной эволюции и времени расхождения в отсутствие молекулярных часов
,
Биоинформатика
,
2003
, том.
19
(стр.
301
—
302
)
Schloss
P.D.
,
Handelsman
J.
.
К переписи бактерий в почве
,
PLoS Comp. биол.
,
2006
, том.
2
(стр.
786
—
793
)
Шаффер
Х.Б.
,
Томсон
Р.К.
.
Разграничение видов в недавних излучениях
,
Сист. биол.
,
2007
, том.
56
(стр.
896
—
906
)
Сайты
J.W.
,
Marshall
J.C.
.
Разграничение видов: проблема возрождения систематической биологии
,
Trends Ecol. Эвол.
,
2003
, том.
18
(стр.
462
—
470
)
Стаматакис
А.2 А.
RAxML-VI-HPC: филогенетический анализ на основе максимального правдоподобия с использованием тысяч таксонов и смешанных моделей
22
(стр.
2688
—
2690
)
Тан
C.Q.
,
Leasi
F.
,
Obertegger
U.
,
Kieneke
A.
,
Barraclough
T.G.G.G.G.G.
,
T.G.G.G.G.
,
T.G.
,
.
,
Фонтането
Д.
.
Широко используемая молекула малой субъединицы 18S рДНК сильно недооценивает истинное разнообразие в исследованиях биоразнообразия мейофауны
,
Проц. Натл. акад. науч. США
,
2012
, том.
109
(стр.
16208
—
16212
)
Templeton
3
Отте
Д.
,
Эндлер
Дж.
.
Значение видов и видообразования: популяционно-генетический подход
,
Видообразование и его последствия
,
1989
Сандерленд (Массачусетс)
Sinauer Associates
Темплтон
А. Р.
.
Использование филогеографического анализа деревьев генов для проверки статуса видов и процессов
,
Mol. Экол.
,
2001
, том.
10
(стр.
779
—
791
)
Venter
20 J.C.
002 Remington
K.
,
Heidelberg
J.F.
,
HALPERN
A.L.
,
. 9000.222,
. 9000.222,
. 9000. 9000. 9000. 9000... 9000.. 9000.22.
,
,
Полсен
I.
,
Нельсон
К.Е.
,
Нельсон
В.
,
Фаутс
Д.Е.
,
Levy
S.
,
Knap
A.H.
,
Lomas
M.W.
,
Nealson
K.
,
White
O.
,
Peterson
J.
,
Hoffman
J.
,
Parsons
R.
,
Baden-Tillson
H.
,
Pfanls0003
К.
,
Роджерс
Ю. Х.
,
Смит
Х.О.
.
Секвенирование экологического генома Саргассова моря с дробовиком
304
(стр.
66
—
74
)
Wiens
J.J.
,
Пенкрот
Т.А.
.
Разграничение видов с использованием ДНК и морфологической изменчивости и дискордантных пределов видов у шипастых ящериц ( Sceloporus )
,
Syst. биол.
,
2002
, том.
51
(стр.
69
—
91
)
Воля
КВт.
,
Рубинофф
Д.
.
Миф о молекуле: штрих-коды ДНК для видов не могут заменить морфологию для идентификации и классификации
,
Кладистика
,
2004
, том.
20
(стр.
47
—
55
)
Ян
Z.
3 9. ,
вычислительная молекулярная эволюция
,
2006
Оксфорд
Oxford University Press
Ян
Z.
,
Goldman
N.
2,
пятница
N.
,
N.2,
N.
,
N.
,
.0003
А.
.
Деревья максимального правдоподобия из последовательностей ДНК: своеобразная задача статистической оценки
,
Сист. биол.
,
1995
, том.
44
(стр.
384
—
399
)
Ян
Z.
,
Rannala
B.
22222.
Байесовское разграничение видов с использованием данных о многолокусных последовательностях
,
Проц. Натл. акад. науч. США
,
2010
, том.
107
(стр.
9264
—
9269
)
Автор. Примечания
Ассоциированный редактор: Ричард Глор
© The Poort (S) 2013. Общество систематических биологов.
Это статья в открытом доступе, распространяемая в соответствии с лицензией Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), которая разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии, что оригинальная работа правильно цитируется.
© Автор(ы), 2013 г. Опубликовано Oxford University Press от имени Общества систематических биологов.
Раздел выпуска:
Обычные статьи
Скачать все слайды
Дополнительные данные
Дополнительные данные
AddSuppFiles-2 — файл html
AddSuppFiles-1 — файл docx
Реклама
Цитаты
Просмотры
13 855
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи через
Последний
Самые читаемые
Самые цитируемые
Сексуальные сигналы сохраняются в течение длительного времени: древняя кооптация биолюминесценции для демонстрации ухаживания у карпообразных остракод
Важность контактных зон для различения межвидовой и внутривидовой географической изменчивости
Прогнозирование длинных висячих краев в модельных филогениях с приложениями к биоразнообразию и выводу деревьев
Функциональные компромиссы асимметрично способствуют фенотипической эволюции
Объявления
Реклама
900:00 ЮЛЬ — 21 декабря 2022 г. Йоль наступает 21 декабря, в день зимнего солнцестояния. Это самый короткий день и самая длинная ночь в году, так как Солнце находится на самой низкой высоте в небе. Помимо обозначения нового сезона, зимнее солнцестояние также является важным символом Солнца. Кажется, что в этот день оно стоит на месте, считается, что Солнце переживает перерождение. Затем он будет набирать обороты после солнцестояния. Праздник также изображает несколько языческих верований, особенно верование Короля Остролиста, передающего власть Королю Дубов, открывая новый сезон света.
История Йоля
Наши предки зависели от течения времен и сезонов. И лучший способ измерить времена года — это наблюдать за Солнцем и орбитой Земли вокруг него. Зимнее солнцестояние — это время года, когда Солнце возрождается, объявляя о новом сезоне.
21 декабря день короче, так как кажется, что Солнце стоит на более низкой высоте, из-за чего ночь становится длиннее. Но это переходный период, который открывает новый сезон большего количества солнечного света. Празднование Йоля раньше было связано с различными языческими традициями, граничащими с мифологией и культурой. Популярные представления включают миф о богине, родившей бога Солнца. Существует также празднование передачи власти от Короля Остролиста Королю-Дубу. Плюс древний праздник германцев о Дикой Охоте и боге Одине тоже примерно в это же время.
Все вышеперечисленное составляет основу празднования Йоля. Но, как было сказано ранее, она еще более символична, поскольку помогала нашим предкам определять времена и времена года. Понимание времен и сезонов обычно было разницей между жизнью и смертью, доступностью пищи и голодом, победой и поражением и многими другими вещами. С введением христианства празднование Йоля также было связано с рождественскими традициями. Считается, что 25 декабря было выбрано для празднования рождения Христа, потому что это период обновления. Йоль и Рождество имеют определенное сходство, и они также часто пересекаются.
Хронология Йоля
4 г. н.э.
Ранние отчеты о праздновании
Йоль отмечается уже в 4 г. н.э. немецкими готами.
726 г. н.э.
Упоминается старое написание Йоля
Св. Беда упоминает «Джули», изображая праздничные дни с декабря по январь.
1475
Самое раннее употребление слова Юлетид
Существительное Юлетид используется в период Рождества.
\\1500
Лучшее управление продовольствием
Постоянный голод, который обычно преследовал зимние месяцы, сокращен.
Йоль
Часто задаваемые вопросы s Является ли Йоль таким же, как Рождество?
Хотя празднования часто совпадают, это два разных праздника.
Что такое Святки?
Это период вокруг Рождества, с 21 декабря по 1 января.
Является ли Йоль языческим праздником?
Праздник имеет некоторую историю языческих традиций, но современные праздники могут не учитывать их.
Святочные мероприятия
Украсьте свой дом
Во время Святочного праздника вы можете украсить свой дом Святочными артефактами. К ним относятся вечнозеленые деревья, венки, ветки, свечи и многое другое.
Приготовьте традиционное Святочное блюдо
Некоторые блюда характерны для празднования Йоля, и их легко приготовить. Примером может служить очарование хлеба на пахте.
Дарите подарки и празднуйте с друзьями
Во время праздника Йоль вы можете дарить подарки друзьям. А также собраться у костра, чтобы отпраздновать возрождение света.
5 Интересные факты о зимнем солнцестоянии
Это происходит два раза в год
Зимнее солнцестояние происходит два раза в год, один раз в Южном полушарии, а также в Северном.
Самая длинная ночь в году
Это день, который сочетается с самой длинной ночью.
Самый короткий день в году
Кстати, в день зимнего солнцестояния Солнце появляется на очень короткий период.
Значение солнцестояния
Солнцестояние — это сочетание двух латинских слов, означающих «Солнце стоит на месте».
Летнее солнцестояние в июне
В то время как зимнее солнцестояние происходит в декабре, противоположное явление происходит в июне.
Почему мы любим Йоль
Это дает лучшее понимание времен года
Во время празднования Йоля мы лучше понимаем наше Солнце. Мы лучше понимаем смену времен года и орбиту нашей Земли.
Это праздник света
Зимнее солнцестояние знаменует возрождение Солнца. Таким образом, он считается праздником света.
Он богат историей
В праздник Йоль мы узнаем больше о наших предках. В частности, мы также можем понять их деятельность в зимние периоды.
Святочные даты
Год Дата День 2022 December 21 Wednesday 2023 December 21 Thursday 2024 December 21 Saturday 2025 December 21 Sunday 2026 21 декабря Понедельник
Лучшие видеоролики Yule Log на YouTube, от Sonic до Marvel и Minecraft
Видеоролики из рождественского журнала стали праздничным увлечением и небольшим ответвлением культуры потокового вещания. Их легче собрать, чем настоящий костер, они более практичны для жителей квартир или людей, живущих в теплом климате, и не требуют очистки. Найти его в Интернете — это быстрый и простой процесс, и они придают согревающий цвет и праздничный тон для праздничных торжеств. Но поскольку их также легко создавать и распространять, внезапно у каждой потоковой платформы появляются собственные каминные видео.
И так, кажется, в каждой развлекательной франшизе. Есть два официальных журнала Frozen Yule Logs, несколько журналов Marvel и фирменные журналы Yule Logs для Hearthstone и Minecraft , Outlander и Shrek . Нанесение названия франшизы и темы на видео у камина — это забава и игра для крупных развлекательных корпораций, пока вы не остановитесь на мгновение и не подумаете: «Подождите, значит ли это, что это Рождественское полено — канон?!»
Хороший вопрос. К праздничному сезону 2021 года мы обновили наш исчерпывающий обзор видеороликов «Рождественский журнал», посвященных франшизе, из Интернета, оценили их фактор удовольствия, а также их более глубокие канонические последствия и отобрали лучшие. Нам здесь весело .
Святочное бревно замка Эренделл (Disney Plus) Изображение: Walt Disney Pictures Замороженные святочные бревна (Disney Plus): Святочное бревно замка Эренделл и Святочное бревно замка Эренделл: бумажное издание
Как долго они? Около трех часов каждый, идеально подходит для фоновой игры во время Family Game Night.
Уровни рождественского настроения? Святочное полено в замке Эренделл может похвастаться подарками, бережно развешенными у огня чулками и крошечной рождественской елкой сбоку. Есть гирлянды и остролист, все это создает ощущение уютного праздника. Никакой праздничной музыки или каверов на «Let it Go», так что это исключительно каминный опыт.
Tte Paper Edition включает в себя вырезанные изображения Эльзы, Анны, Кристоффа, Олафа и Свена, а также языки пламени из бумаги. Это довольно красиво, хотя и не специально для Рождества.
Подходят ли к Canon? Это тот самый камин, перед которым Эльза, Анна, Кристофф, Олаф и Свен играют в шарады в Frozen 2 . Эренделл канонически празднует множество зимних праздников, как видно из 21-минутного короткометражного фильма «Замороженное приключение Олафа», поэтому этот сеттинг Рождественского журнала идеально вписывается в мир.
Бумажное издание на самом деле не имеет никакой канонической ценности, но кто сказал, что в мире Frozen существует , а не ? .
Домашний камин: Bright Edition (Netflix) Изображение: Netflix Сколько времени? Оттенок выше часа.
Уровни рождественского настроения? Если вы называете песчаный, красочный, буквально мусорный огонь рождественским весельем, то в нем его много.
Подходит ли к Canon? Да. Граффити на стене и мусорный бак — все это элементы, которые идеально вписываются в безобразный уличный мир Bright 9. 0094, который является «кинематографическим воплощением лопнувшей канализационной трубы».
DC Yole Log Сколько времени? 10 часов.
Уровни рождественского настроения? Вся сцена празднична и украшена чулками над очагом, деревом в углу и менорой на столе.
Подходит ли к Canon? Есть чулки для Дианы, Кларка, Брюса и Дика (которые в конце меняются на Джейсона). Трезубец покоится на камине, и вместо мишуры это, кажется, лассо Чудо-женщины, украшающее очаг. Это канон? Трудно сказать, зачем там камин, раскрывающий кучу тайных личностей, но это мило.
Дэдпул Йол Бревно Сколько времени? Всего пять минут.
Уровни рождественского настроения? Буквальный горящий бумажный пакет. Но он установлен на празднично украшенном пороге, и на грубом рисунке Дэдпула действительно есть шапка Санты.
Подходит ли к Canon? Конечно. Тотализатор, где делаются ставки на смерть знаменитостей. Смешной.
ТАРДИС Святочное бревно | Доктор Кто Рождество | Рождественская ночь Журнал Праздничного Тринадцатого Доктора Юла Сколько времени? Первый чуть больше двух часов, второй почти ровно два часа.
Уровни рождественского настроения? У первого на заднем плане рождественский камин с чулками, гирляндой и елкой. Однако центральным элементом является снежный шар ТАРДИС с небольшим венком внутри. В этом также есть прекрасная оркестровая музыка, оригинальная для шоу. Это не совсем рождественское настроение, но вместо этого оно очень похоже на 9.0093 Доктор Кто в тон.
Второй — полностью украшенный камин с чулками, гирляндой ТАРДИС, огнями и деревом. Наверху висят гирлянды, а когда камера поворачивается, мы видим тарелку с печеньем. (Э-э, печенье, поскольку это британское видео.) Как мило! Но примечательно, что за окном будто падают листья, а не снег.
Подходит ли к Canon? В первом видео есть несколько странных моментов, когда Доктора выходят из снежного шара ТАРДИС, и есть реклама 2017 Доктор Кто Рождественский спецвыпуск, так что действие явно происходит не в сериале.
Второй, однако, более четко соответствует канону. Несколько раз появляется Тринадцатый Доктор, чтобы согреть ей руки. В какой-то момент она улетает в ТАРДИС, а затем возвращается, чтобы снова разжечь огонь. Мы не знаем, где это или что она делает, но, учитывая, что вся фишка Доктора заключается в возможности путешествовать во времени и пространстве, это может сработать где угодно и когда угодно.
Американская история ужасов и что мы делаем в тени Святочные журналы На официальном канале FX есть подборка того, что я могу назвать только ироничными Святочными журналами. Они праздничные? Нет. Они абсолютно веселые? Да. Они, безусловно, заслуживают включения, но, возможно, не допрашиваются на предмет их канонической ценности или уровня одобрения. Оба длятся около получаса и фактически органично вписываются в уже существующие сцены.
Девушка с моря Рождественское бревно Сколько времени? Всего 10 минут.
Уровни рождественского настроения? Если вы не из Флориды или другого места, тепло не обязательно означает праздничное настроение.
Подходит ли к Canon? Сцена в видео выглядит так, как будто она идеально подходит для графического романа Молли Нокс Остертаг. Это может быть не супер-Рождество, но это очень мило и достойно включения.
Святочное бревно Аида Сколько времени? Колоссальные 12 часов. Вы можете держать это на протяжении всего Рождества.
Уровни рождественского настроения? Не так уж и много, учитывая, что действие Аида происходит не только до популяризации современных декоративных пробных камней Рождества, но и до появления христианства. Тем не менее, есть гранатовые венки и большой старый мешок сатира, так что это очень празднично в духе Подземного мира!
Подходит ли к Canon? Ага! Все пробные камни на месте, и почти ожидаешь, что Цербер подойдет и уютно вздремнет у камина.
Святочное бревно мурлока Hearthstone Сколько времени? Волосы более 53 минут, хороши в качестве фона открытия подарка.
Уровни рождественского настроения? Поначалу не так много, но чем больше мурлоков появляется на экране, тем больше и больше украшается камин. К концу видео это полностью празднично украшенная сцена. Время от времени фоном играет рождественская музыка, но ее поют мурлоки.
Подходит ли к канону? Действие происходит в таверне, где предположительно происходит Hearthstone . Вы можете услышать звук людей, играющих в игру. Декор посвящен миру, как и вышеупомянутая музыка в исполнении мурлоков. В целом, это очаровательное дополнение к канону Hearthstone .
Камины Marvel Comics Есть несколько каминов, характерных для комиксов Marvel, но некоторые из них, в частности, , а не , вызывают рождественское настроение (серьезно, одинокая, безрадостная квартира Тони Старка просто унылая).
Видео у камина в квартире Капитана Америки в Бруклине в 4K Сколько времени? Несколько секунд больше часа.
Уровни рождественского настроения? Квартира Кэпа довольно пуста, но на заднем плане играют джазовые рождественские стандарты, чтобы задать тон. Это не так тепло, как , как болтовня толпы, но все же это что-то.
Подходит ли к Canon? В большей степени, чем в некоторых других рождественских бревнах кинематографической вселенной Marvel. Версия крупным планом показывает обрамленное изображение бывшего любовного интереса Пегги, что трогательно, хотя и немного грустно, поскольку вся эта серия у камина, похоже, происходит в стихах комиксов. Даже если это не так, это видео 2016 года, так что оно до Мстители: Финал Воссоединение Стегги.
Домашний камин мисс Марвел в Нью-Джерси в формате 4K (крупный план)
Сколько времени? Час и сдача.
Сколько рождественского настроения? Немного, но это нормально, потому что Камала Хан не празднует Рождество. Это теплый камин холодной зимой в Нью-Джерси и хорошее время, чтобы насладиться неконфессиональным теплом. Мы слышим болтовню на вечеринке, что само по себе приносит некоторую праздничную радость.
Подходит ли к Canon? Нет ничего, что указывало бы на то, что это канон, но и ничего, что указывало бы на то, что это не так. Маленькая милая деталь — костюм Камалы, накинутый на ее диван — лучше спрячьте его, пока кто-нибудь не увидел!
Майнкрафт Праздничный журнал Йоля Сколько времени? Два часа, то есть как раз в середине большинства этих видео Йольского журнала. Недостаточно долго, чтобы развлекать вас весь день, но и не слишком коротко, чтобы вам пришлось слишком много раз нажимать «перезагрузка».
Уровни рождественского настроения? Так много! Очаг украшен, на улице идет снег, подарков в изобилии. Такой уютный, такой необычный. Это идеальный рождественский опыт. Время от времени на экране появляются приятные рождественские сюрпризы.
Подходит ли к Canon? Minecraft канон — это то, что вы делаете из него, так что, конечно, это может быть каноном. Может быть, нет настоящего NPC Санты, но может быть , если вы очень верите, что мы имеем в виду, если вы загружаете скин Санты. Также в какой-то момент кто-то врывается в дом и крадет подарок. Разве это не Майнкрафт способ?
Чужеземец | Святочное бревно (2014) Чужестранка | Кухня Фрейзера ![]() com/embed/7S8It-zl63s?rel=0″ allowfullscreen=»» scrolling=»no» allow=»encrypted-media; accelerometer; gyroscope; picture-in-picture»> Чужестранка | Журнал Fraser’s Ridge Yole Log (2019)
com/embed/7S8It-zl63s?rel=0″ allowfullscreen=»» scrolling=»no» allow=»encrypted-media; accelerometer; gyroscope; picture-in-picture»> Чужестранка | Журнал Fraser’s Ridge Yole Log (2019) Как долго это длится? Рождественская версия 2019 и 2014 года длится около часа. Кухня 2019 года — около 90 минут.
Уровни рождественского настроения? Более тонкий декор, соответствующий старинным предметам. Два из них содержат животных, которые появляются в шоу, которое не совсем рождественское, но очень веселое. Двое из них играют главную музыкальную тему, которая на самом деле звучит довольно по-рождественски, в грустной манере, как в «O Holy Night». Затем он переключается на общую праздничную кельтскую музыку.
Кухня — исключение, здесь только горящий камин и минимальное праздничное убранство.
Подходит ли к Canon? Включение кота в последнее видео — приятный штрих, чтобы связать эти видео с камином с каноном. В более свежем видео есть котенок Адсо, которого представили в последнем сезоне Outlander . У того, что было пять лет назад, есть две собаки, у которых на самом деле нет никаких канонических связей, но они, безусловно, милые! В целом, они стилистически вписываются в сериал, и этого достаточно.
Seinfeld Festivus Yole Log Сколько времени? Чуть больше часа.
Уровни рождественского настроения? Этому видео явно не хватает рождественского настроения, но это нормально, потому что оно полно праздничного настроения!
Подходит ли к Canon? Не уверен, чья это квартира, или у кого из команды Сайнфелда вообще был бы такой роскошный камин в их квартирах, но точно! Единственное, что действительно выбивает его из канона, — это крошечная книжечка с надписью «Сайнфелд» в корзине.
Журнал Шрека Йола Сколько времени? Чуть меньше 30 минут, так что вам может понадобиться нажать кнопку повтора пару раз.
Уровни рождественского настроения? Сам очаг украшен гирляндами. На фоне играют какие-то веселые мелодии. Слышно болтовню детей и гостей. Чего действительно не хватает, так это звука огня, который теряется под персонажами Шрека, время от времени блуждающими по экрану.
Подходит ли к Canon? В короткометражном спин-оффе Shrek the Halls установлено, что Рождество действительно существует во вселенной Шрека, и короткометражка о том, что это Рождественское бревно изначально было соединено с Рождественским шректакуляром Ослика , продолжает эту тенденцию. Мы сразу видим, где мы находимся — болотная гавань Шрека — и куда вписывается эта сцена: Шрек зажигает огонь прямо перед тем, как он и Фиона пригласят гостей на праздники. Мы — это во вселенной Шрека, тусуемся со Шреком и его семьей. Идеальные часы, если вы жаждете теплого семейного общения, но у вас есть только YouTube.
Как приручить дракона: журнал Snoggletog Сколько времени? Также немного меньше 30 минут, так что, как и в случае с Святочным журналом Шрека, вам нужно будет нажать повтор. Или играть их спина к спине!
Уровни рождественского настроения? «Бревно сноглетог» — это просто окно в фантастическое Рождество в мире Как приручить дракона , вплоть до украшений и веселой рождественской музыки, играющей на заднем плане. Как и в случае с видео Шрека, многие настоящие звуки огня теряются среди фоновой музыки и персонажей, блуждающих по экрану.
Подходит ли к Canon? Ага! Подобно тому, как Шрек был компаньоном «Рождественский шректакуляр осла», «Бревно сноглетог» является компаньоном «Как приручить дракона: Возвращение домой », премьера которого состоялась ранее в этом месяце на канале NBC. Действие короткометражного фильма происходит после событий финального фильма «Как приручить дракона », в котором исследуются последствия трилогии. У Иккинга и Астрид есть дети, а также у Беззубика и Дневной Фурии, и все они появляются здесь. Вполне вписывается в канон.
Sonic 2 Yule Log Сколько времени? Час!
Уровень рождественского настроения? Все начинается довольно скромно, но по ходу видео Соник бежит и получает все больше и больше декораций, пока весь очаг не становится невероятно праздничным. Там есть елка, чулки, подарки, список желаний для Санты и многое другое.
Подходит ли к Canon? У главных героев есть собственные чулки, и Соник, вероятно, из тех парней, которые очень рады праздникам.
Venom Yole Logs В настоящее время их два: симпатичный анимированный и более реалистичный для Venom: Let There Be Carnage .
Сколько времени? Первая длится 10 часов, но кажется, что это всего лишь 10-секундный цикл анимированного Венома, который слегка подпрыгивает и пьет горячее какао. (Отказ от ответственности: мы не смотрели все 10 часов. Может быть, он делает что-то сложное и увлекательное за семь часов.)
Второй также длится 10 часов, при этом огни меняются и мигают через определенные промежутки времени.
Уровни рождественского настроения? Анимированный Веном сидит рядом с уютным камином под портретом Венома, одетого в причудливый шелковый жакет богатого человека.
П.92
92
92
92
APE: анализ филогенетики и эволюции на языке R
,
Биоинформатика
,
2004
, том.
20
(стр.
289
—
290
)
Phillimore
A.B.
,
Цена
Т.Д.
.
Зависимый от плотности кладогенез у птиц
,
PLoS Biol.
,
2008
, том.
6
стр.
e71
Pons
J.
,
Барраклаф
Т.Г.
,
Гомес-Зурита
Дж.
,
Кардосо
А.
,
Дюран
3
,
Hazell
S.
,
Kamoun
S.
,
Sumlin
W.D.
,
Vogler
A.P.
,
A.P.
,
A. P.
,
A.D.
Разграничение видов на основе последовательностей для таксономии ДНК неописанных насекомых
,
Сист. биол.
,
2006
, том.
55
(pg.
595
—
609
)
Pons
J.
,
Fujisawa
T.
,
Claridge
E.M.
,
Сэвилл
Р.А.
,
Барраклаф
Т.Г.
,
Фоглер
А. П.
.
Глубокое подразделение мтДНК у линнеевских видов в эндемической радиации тигровых жуков из Новой Зеландии (род Neocicindela) Фил. Эвол.
,
2011
, том.
59
(стр.
251
—
262
)
Пауэлл
2Дж. Р. 9003.
Учет неопределенности в разграничении видов при анализе данных о последовательностях ДНК из окружающей среды
,
Методы Экол. Эвол.
,
2012
, том.
3
(стр.
1
—
11
)
Pybus
O.G.
,
Харви
П.Х.
.
Тестирование макроэволюционных моделей с использованием неполных молекулярных филогений
,
Proc. Р. Соц. Лонд. Б
,
2000
, том.
267
(стр.
2267
—
2272
)
R Development Core Team
,
R: A Язык и окружающая среда для статистических вычислений
,
20102 2010 2010 20102 2010 20102 2010 2010 20102 2010 2010 20102 20102 2010 2010 20102 2010 20102 20102 2010 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 2010 20102 20102 2010 2010 2 20102 20102 2010 2010 2 20102 2010 2010 2 20102 2010 2010 2010 2 2010 2 20102 2010 2010 2010 2 20102. )
R Фонд статистических вычислений
Рабоски
Д.Л.
,
Ловетт
И.Дж.
.
Зависимая от плотности диверсификация североамериканских лесных камышевок
,
Проц. Р. Соц. Лонд. Б
,
2008
, том.
275
(стр.
2363
—
2371
)
Рамбо
А.
Phylo-O-Gen: Phylogenetic Tree Simulator Package v1.1
,
2002
Rambaut
A.
,
Gravly
N.C.
222222222222222.
Seq-Gen: приложение для моделирования эволюции последовательностей ДНК вдоль филогенетических деревьев методом Монте-Карло
,
CABIOS
,
1997
, том.
13
(стр.
235
—
238
)
Раннала
B.
,
Yang
Z.
2.
Байесовская оценка времени дивергенции видов и размера предковой популяции с использованием последовательностей ДНК из нескольких локусов
,
Генетика0222 , том.
164
(стр.
1645
—
1656
)
Reid
N.M.
,
Carstens
г. до н.э.
.
Ошибка филогенетической оценки может снизить точность разграничения видов: байесовская реализация модели General Mixed Yule Coalescent
,
BMC Evol. биол.
,
2012
, том.
12
стр.
196
Риклефс
Р.Э.
.
Оценка темпов диверсификации на основе филогенетической информации
,
Trends Ecol. Эвол.
,
2007
, том.
22
(стр.
601
—
610
)
Rosenberg
N.A.
,
Nordborg
M. 9000
2222.
Генеалогические деревья, теория слияния и анализ генетических полиморфизмов
,
Нац. Преподобный Жене.
,
2002
, том.
3
(стр.
380
—
390
)
Sanderson 2
M.J.
3.
r8s: вывод абсолютных скоростей молекулярной эволюции и времени расхождения в отсутствие молекулярных часов
,
Биоинформатика
,
2003
, том.
19
(стр.
301
—
302
)
Schloss
P.D.
,
Handelsman
J.
.
К переписи бактерий в почве
,
PLoS Comp. биол.
,
2006
, том.
2
(стр.
786
—
793
)
Шаффер
Х.Б.
,
Томсон
Р.К.
.
Разграничение видов в недавних излучениях
,
Сист. биол.
,
2007
, том.
56
(стр.
896
—
906
)
Сайты
J.W.
,
Marshall
J.C.
.
Разграничение видов: проблема возрождения систематической биологии
,
Trends Ecol. Эвол.
,
2003
, том.
18
(стр.
462
—
470
)
Стаматакис
А.2 А.
RAxML-VI-HPC: филогенетический анализ на основе максимального правдоподобия с использованием тысяч таксонов и смешанных моделей
22
(стр.
2688
—
2690
)
Тан
C.Q.
,
Leasi
F.
,
Obertegger
U.
,
Kieneke
A.
,
Barraclough
T.G.G.G.G.G.
,
T.G.G.G.G.
,
T.G.
,
.
,
Фонтането
Д.
.
Широко используемая молекула малой субъединицы 18S рДНК сильно недооценивает истинное разнообразие в исследованиях биоразнообразия мейофауны
,
Проц. Натл. акад. науч. США
,
2012
, том.
109
(стр.
16208
—
16212
)
Templeton
3
Отте
Д.
,
Эндлер
Дж.
.
Значение видов и видообразования: популяционно-генетический подход
,
Видообразование и его последствия
,
1989
Сандерленд (Массачусетс)
Sinauer Associates
Темплтон
А. Р.
.
Использование филогеографического анализа деревьев генов для проверки статуса видов и процессов
,
Mol. Экол.
,
2001
, том.
10
(стр.
779
—
791
)
Venter
20J.C.
002 Remington
K.
,
Heidelberg
J.F.
,
HALPERN
A.L.
,
. 9000.222,
. 9000.222,
. 9000. 9000. 9000. 9000... 9000.. 9000.22.
,
,
Полсен
I.
,
Нельсон
К.Е.
,
Нельсон
В.
,
Фаутс
Д.Е.
,
Levy
S.
,
Knap
A.H.
,
Lomas
M.W.
,
Nealson
K.
,
White
O.
,
Peterson
J.
,
Hoffman
J.
,
Parsons
R.
,
Baden-Tillson
H.
,
Pfanls0003
К.
,
Роджерс
Ю. Х.
,
Смит
Х.О.
.
Секвенирование экологического генома Саргассова моря с дробовиком
304
(стр.
66
—
74
)
Wiens
J.J.
,
Пенкрот
Т.А.
.
Разграничение видов с использованием ДНК и морфологической изменчивости и дискордантных пределов видов у шипастых ящериц ( Sceloporus )
,
Syst. биол.
,
2002
, том.
51
(стр.
69
—
91
)
Воля
КВт.
,
Рубинофф
Д.
.
Миф о молекуле: штрих-коды ДНК для видов не могут заменить морфологию для идентификации и классификации
,
Кладистика
,
2004
, том.
20
(стр.
47
—
55
)
Ян
Z.
3 9. ,
вычислительная молекулярная эволюция
,
2006
Оксфорд
Oxford University Press
Ян
Z.
,
Goldman
N.
2,
пятница
N.
,
N.2,
N.
,
N.
,
.0003
А.
.
Деревья максимального правдоподобия из последовательностей ДНК: своеобразная задача статистической оценки
,
Сист. биол.
,
1995
, том.
44
(стр.
384
—
399
)
Ян
Z.
,
Rannala
B.
22222.
Байесовское разграничение видов с использованием данных о многолокусных последовательностях
,
Проц. Натл. акад. науч. США
,
2010
, том.
107
(стр.
9264
—
9269
)
Автор. Примечания
Ассоциированный редактор: Ричард Глор
© The Poort (S) 2013. Общество систематических биологов.
Это статья в открытом доступе, распространяемая в соответствии с лицензией Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), которая разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии, что оригинальная работа правильно цитируется.
© Автор(ы), 2013 г. Опубликовано Oxford University Press от имени Общества систематических биологов.
Раздел выпуска:
Обычные статьи
Скачать все слайды
Дополнительные данные
Дополнительные данные
AddSuppFiles-2 — файл html
AddSuppFiles-1 — файл docx
Реклама
Цитаты
Просмотры
13 855
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи через
Последний
Самые читаемые
Самые цитируемые
Сексуальные сигналы сохраняются в течение длительного времени: древняя кооптация биолюминесценции для демонстрации ухаживания у карпообразных остракод
Важность контактных зон для различения межвидовой и внутривидовой географической изменчивости
Прогнозирование длинных висячих краев в модельных филогениях с приложениями к биоразнообразию и выводу деревьев
Функциональные компромиссы асимметрично способствуют фенотипической эволюции
Объявления
Реклама
900:00 ЮЛЬ — 21 декабря 2022 г.Йоль наступает 21 декабря, в день зимнего солнцестояния. Это самый короткий день и самая длинная ночь в году, так как Солнце находится на самой низкой высоте в небе. Помимо обозначения нового сезона, зимнее солнцестояние также является важным символом Солнца. Кажется, что в этот день оно стоит на месте, считается, что Солнце переживает перерождение. Затем он будет набирать обороты после солнцестояния. Праздник также изображает несколько языческих верований, особенно верование Короля Остролиста, передающего власть Королю Дубов, открывая новый сезон света.
История Йоля
Наши предки зависели от течения времен и сезонов. И лучший способ измерить времена года — это наблюдать за Солнцем и орбитой Земли вокруг него. Зимнее солнцестояние — это время года, когда Солнце возрождается, объявляя о новом сезоне.
21 декабря день короче, так как кажется, что Солнце стоит на более низкой высоте, из-за чего ночь становится длиннее. Но это переходный период, который открывает новый сезон большего количества солнечного света. Празднование Йоля раньше было связано с различными языческими традициями, граничащими с мифологией и культурой. Популярные представления включают миф о богине, родившей бога Солнца. Существует также празднование передачи власти от Короля Остролиста Королю-Дубу. Плюс древний праздник германцев о Дикой Охоте и боге Одине тоже примерно в это же время.
Все вышеперечисленное составляет основу празднования Йоля. Но, как было сказано ранее, она еще более символична, поскольку помогала нашим предкам определять времена и времена года. Понимание времен и сезонов обычно было разницей между жизнью и смертью, доступностью пищи и голодом, победой и поражением и многими другими вещами. С введением христианства празднование Йоля также было связано с рождественскими традициями. Считается, что 25 декабря было выбрано для празднования рождения Христа, потому что это период обновления. Йоль и Рождество имеют определенное сходство, и они также часто пересекаются.
Хронология Йоля
4 г. н.э.
Ранние отчеты о праздновании
Йоль отмечается уже в 4 г. н.э. немецкими готами.
726 г. н.э.
Упоминается старое написание Йоля
Св. Беда упоминает «Джули», изображая праздничные дни с декабря по январь.
1475
Самое раннее употребление слова Юлетид
Существительное Юлетид используется в период Рождества.
\\1500
Лучшее управление продовольствием
Постоянный голод, который обычно преследовал зимние месяцы, сокращен.
Йоль
Часто задаваемые вопросы sЯвляется ли Йоль таким же, как Рождество?
Хотя празднования часто совпадают, это два разных праздника.
Что такое Святки?
Это период вокруг Рождества, с 21 декабря по 1 января.
Является ли Йоль языческим праздником?
Праздник имеет некоторую историю языческих традиций, но современные праздники могут не учитывать их.
Святочные мероприятия
Украсьте свой дом
Во время Святочного праздника вы можете украсить свой дом Святочными артефактами.
К ним относятся вечнозеленые деревья, венки, ветки, свечи и многое другое.Приготовьте традиционное Святочное блюдо
Некоторые блюда характерны для празднования Йоля, и их легко приготовить. Примером может служить очарование хлеба на пахте.
Дарите подарки и празднуйте с друзьями
Во время праздника Йоль вы можете дарить подарки друзьям. А также собраться у костра, чтобы отпраздновать возрождение света.
5 Интересные факты о зимнем солнцестоянии
Это происходит два раза в год
Зимнее солнцестояние происходит два раза в год, один раз в Южном полушарии, а также в Северном.
Самая длинная ночь в году
Это день, который сочетается с самой длинной ночью.
Самый короткий день в году
Кстати, в день зимнего солнцестояния Солнце появляется на очень короткий период.
Значение солнцестояния
Солнцестояние — это сочетание двух латинских слов, означающих «Солнце стоит на месте».
Летнее солнцестояние в июне
В то время как зимнее солнцестояние происходит в декабре, противоположное явление происходит в июне.
Почему мы любим Йоль
Это дает лучшее понимание времен года
Во время празднования Йоля мы лучше понимаем наше Солнце. Мы лучше понимаем смену времен года и орбиту нашей Земли.
Это праздник света
Зимнее солнцестояние знаменует возрождение Солнца. Таким образом, он считается праздником света.
Он богат историей
В праздник Йоль мы узнаем больше о наших предках. В частности, мы также можем понять их деятельность в зимние периоды.
Святочные даты
| Год | Дата | День |
|---|---|---|
| 2022 | December 21 | Wednesday |
| 2023 | December 21 | Thursday |
| 2024 | December 21 | Saturday |
| 2025 | December 21 | Sunday |
| 2026 | 21 декабря | Понедельник |
Лучшие видеоролики Yule Log на YouTube, от Sonic до Marvel и Minecraft
Видеоролики из рождественского журнала стали праздничным увлечением и небольшим ответвлением культуры потокового вещания. Их легче собрать, чем настоящий костер, они более практичны для жителей квартир или людей, живущих в теплом климате, и не требуют очистки. Найти его в Интернете — это быстрый и простой процесс, и они придают согревающий цвет и праздничный тон для праздничных торжеств. Но поскольку их также легко создавать и распространять, внезапно у каждой потоковой платформы появляются собственные каминные видео.
И так, кажется, в каждой развлекательной франшизе. Есть два официальных журнала Frozen Yule Logs, несколько журналов Marvel и фирменные журналы Yule Logs для Hearthstone и Minecraft , Outlander и Shrek . Нанесение названия франшизы и темы на видео у камина — это забава и игра для крупных развлекательных корпораций, пока вы не остановитесь на мгновение и не подумаете: «Подождите, значит ли это, что это Рождественское полено — канон?!»
Хороший вопрос. К праздничному сезону 2021 года мы обновили наш исчерпывающий обзор видеороликов «Рождественский журнал», посвященных франшизе, из Интернета, оценили их фактор удовольствия, а также их более глубокие канонические последствия и отобрали лучшие. Нам здесь весело .
Святочное бревно замка Эренделл (Disney Plus) Изображение: Walt Disney Pictures
Замороженные святочные бревна (Disney Plus): Святочное бревно замка Эренделл и Святочное бревно замка Эренделл: бумажное издание
Как долго они? Около трех часов каждый, идеально подходит для фоновой игры во время Family Game Night.
Уровни рождественского настроения? Святочное полено в замке Эренделл может похвастаться подарками, бережно развешенными у огня чулками и крошечной рождественской елкой сбоку. Есть гирлянды и остролист, все это создает ощущение уютного праздника. Никакой праздничной музыки или каверов на «Let it Go», так что это исключительно каминный опыт.
Tte Paper Edition включает в себя вырезанные изображения Эльзы, Анны, Кристоффа, Олафа и Свена, а также языки пламени из бумаги. Это довольно красиво, хотя и не специально для Рождества.
Подходят ли к Canon? Это тот самый камин, перед которым Эльза, Анна, Кристофф, Олаф и Свен играют в шарады в Frozen 2 . Эренделл канонически празднует множество зимних праздников, как видно из 21-минутного короткометражного фильма «Замороженное приключение Олафа», поэтому этот сеттинг Рождественского журнала идеально вписывается в мир.
Бумажное издание на самом деле не имеет никакой канонической ценности, но кто сказал, что в мире Frozen существует , а не ? .
Домашний камин: Bright Edition (Netflix) Изображение: NetflixСколько времени? Оттенок выше часа.
Уровни рождественского настроения? Если вы называете песчаный, красочный, буквально мусорный огонь рождественским весельем, то в нем его много.
Подходит ли к Canon? Да. Граффити на стене и мусорный бак — все это элементы, которые идеально вписываются в безобразный уличный мир Bright 9. 0094, который является «кинематографическим воплощением лопнувшей канализационной трубы».
Сколько времени? 10 часов.
Уровни рождественского настроения? Вся сцена празднична и украшена чулками над очагом, деревом в углу и менорой на столе.
Подходит ли к Canon? Есть чулки для Дианы, Кларка, Брюса и Дика (которые в конце меняются на Джейсона). Трезубец покоится на камине, и вместо мишуры это, кажется, лассо Чудо-женщины, украшающее очаг. Это канон? Трудно сказать, зачем там камин, раскрывающий кучу тайных личностей, но это мило.
Дэдпул Йол Бревно Сколько времени? Всего пять минут.
Уровни рождественского настроения? Буквальный горящий бумажный пакет. Но он установлен на празднично украшенном пороге, и на грубом рисунке Дэдпула действительно есть шапка Санты.
Подходит ли к Canon? Конечно. Тотализатор, где делаются ставки на смерть знаменитостей. Смешной.
ТАРДИС Святочное бревно | Доктор Кто Рождество | Рождественская ночь Журнал Праздничного Тринадцатого Доктора ЮлаСколько времени? Первый чуть больше двух часов, второй почти ровно два часа.
Уровни рождественского настроения? У первого на заднем плане рождественский камин с чулками, гирляндой и елкой. Однако центральным элементом является снежный шар ТАРДИС с небольшим венком внутри. В этом также есть прекрасная оркестровая музыка, оригинальная для шоу. Это не совсем рождественское настроение, но вместо этого оно очень похоже на 9.0093 Доктор Кто в тон.
Второй — полностью украшенный камин с чулками, гирляндой ТАРДИС, огнями и деревом. Наверху висят гирлянды, а когда камера поворачивается, мы видим тарелку с печеньем. (Э-э, печенье, поскольку это британское видео.) Как мило! Но примечательно, что за окном будто падают листья, а не снег.
Подходит ли к Canon? В первом видео есть несколько странных моментов, когда Доктора выходят из снежного шара ТАРДИС, и есть реклама 2017 Доктор Кто Рождественский спецвыпуск, так что действие явно происходит не в сериале.
Второй, однако, более четко соответствует канону. Несколько раз появляется Тринадцатый Доктор, чтобы согреть ей руки. В какой-то момент она улетает в ТАРДИС, а затем возвращается, чтобы снова разжечь огонь. Мы не знаем, где это или что она делает, но, учитывая, что вся фишка Доктора заключается в возможности путешествовать во времени и пространстве, это может сработать где угодно и когда угодно.
На официальном канале FX есть подборка того, что я могу назвать только ироничными Святочными журналами. Они праздничные? Нет. Они абсолютно веселые? Да. Они, безусловно, заслуживают включения, но, возможно, не допрашиваются на предмет их канонической ценности или уровня одобрения. Оба длятся около получаса и фактически органично вписываются в уже существующие сцены.
Сколько времени? Всего 10 минут.
Уровни рождественского настроения? Если вы не из Флориды или другого места, тепло не обязательно означает праздничное настроение.
Подходит ли к Canon? Сцена в видео выглядит так, как будто она идеально подходит для графического романа Молли Нокс Остертаг. Это может быть не супер-Рождество, но это очень мило и достойно включения.
Святочное бревно Аида Сколько времени? Колоссальные 12 часов. Вы можете держать это на протяжении всего Рождества.
Уровни рождественского настроения? Не так уж и много, учитывая, что действие Аида происходит не только до популяризации современных декоративных пробных камней Рождества, но и до появления христианства. Тем не менее, есть гранатовые венки и большой старый мешок сатира, так что это очень празднично в духе Подземного мира!
Подходит ли к Canon? Ага! Все пробные камни на месте, и почти ожидаешь, что Цербер подойдет и уютно вздремнет у камина.
Святочное бревно мурлока HearthstoneСколько времени? Волосы более 53 минут, хороши в качестве фона открытия подарка.
Уровни рождественского настроения? Поначалу не так много, но чем больше мурлоков появляется на экране, тем больше и больше украшается камин. К концу видео это полностью празднично украшенная сцена. Время от времени фоном играет рождественская музыка, но ее поют мурлоки.
Подходит ли к канону? Действие происходит в таверне, где предположительно происходит Hearthstone . Вы можете услышать звук людей, играющих в игру. Декор посвящен миру, как и вышеупомянутая музыка в исполнении мурлоков. В целом, это очаровательное дополнение к канону Hearthstone .
Камины Marvel ComicsЕсть несколько каминов, характерных для комиксов Marvel, но некоторые из них, в частности, , а не , вызывают рождественское настроение (серьезно, одинокая, безрадостная квартира Тони Старка просто унылая).
Видео у камина в квартире Капитана Америки в Бруклине в 4K Сколько времени? Несколько секунд больше часа.
Уровни рождественского настроения? Квартира Кэпа довольно пуста, но на заднем плане играют джазовые рождественские стандарты, чтобы задать тон. Это не так тепло, как , как болтовня толпы, но все же это что-то.
Подходит ли к Canon? В большей степени, чем в некоторых других рождественских бревнах кинематографической вселенной Marvel. Версия крупным планом показывает обрамленное изображение бывшего любовного интереса Пегги, что трогательно, хотя и немного грустно, поскольку вся эта серия у камина, похоже, происходит в стихах комиксов. Даже если это не так, это видео 2016 года, так что оно до Мстители: Финал Воссоединение Стегги.
Домашний камин мисс Марвел в Нью-Джерси в формате 4K(крупный план)
Сколько времени? Час и сдача.
Сколько рождественского настроения? Немного, но это нормально, потому что Камала Хан не празднует Рождество. Это теплый камин холодной зимой в Нью-Джерси и хорошее время, чтобы насладиться неконфессиональным теплом. Мы слышим болтовню на вечеринке, что само по себе приносит некоторую праздничную радость.
Подходит ли к Canon? Нет ничего, что указывало бы на то, что это канон, но и ничего, что указывало бы на то, что это не так. Маленькая милая деталь — костюм Камалы, накинутый на ее диван — лучше спрячьте его, пока кто-нибудь не увидел!
Майнкрафт Праздничный журнал Йоля Сколько времени? Два часа, то есть как раз в середине большинства этих видео Йольского журнала. Недостаточно долго, чтобы развлекать вас весь день, но и не слишком коротко, чтобы вам пришлось слишком много раз нажимать «перезагрузка».
Уровни рождественского настроения? Так много! Очаг украшен, на улице идет снег, подарков в изобилии. Такой уютный, такой необычный. Это идеальный рождественский опыт. Время от времени на экране появляются приятные рождественские сюрпризы.
Подходит ли к Canon? Minecraft канон — это то, что вы делаете из него, так что, конечно, это может быть каноном. Может быть, нет настоящего NPC Санты, но может быть , если вы очень верите, что мы имеем в виду, если вы загружаете скин Санты. Также в какой-то момент кто-то врывается в дом и крадет подарок. Разве это не Майнкрафт способ?
Чужеземец | Святочное бревно (2014) Чужестранка | Кухня ФрейзераКак долго это длится? Рождественская версия 2019 и 2014 года длится около часа. Кухня 2019 года — около 90 минут.
Уровни рождественского настроения? Более тонкий декор, соответствующий старинным предметам. Два из них содержат животных, которые появляются в шоу, которое не совсем рождественское, но очень веселое. Двое из них играют главную музыкальную тему, которая на самом деле звучит довольно по-рождественски, в грустной манере, как в «O Holy Night». Затем он переключается на общую праздничную кельтскую музыку.
Кухня — исключение, здесь только горящий камин и минимальное праздничное убранство.
Подходит ли к Canon? Включение кота в последнее видео — приятный штрих, чтобы связать эти видео с камином с каноном. В более свежем видео есть котенок Адсо, которого представили в последнем сезоне Outlander . У того, что было пять лет назад, есть две собаки, у которых на самом деле нет никаких канонических связей, но они, безусловно, милые! В целом, они стилистически вписываются в сериал, и этого достаточно.
Seinfeld Festivus Yole LogСколько времени? Чуть больше часа.
Уровни рождественского настроения? Этому видео явно не хватает рождественского настроения, но это нормально, потому что оно полно праздничного настроения!
Подходит ли к Canon? Не уверен, чья это квартира, или у кого из команды Сайнфелда вообще был бы такой роскошный камин в их квартирах, но точно! Единственное, что действительно выбивает его из канона, — это крошечная книжечка с надписью «Сайнфелд» в корзине.
Сколько времени? Чуть меньше 30 минут, так что вам может понадобиться нажать кнопку повтора пару раз.
Уровни рождественского настроения? Сам очаг украшен гирляндами. На фоне играют какие-то веселые мелодии. Слышно болтовню детей и гостей. Чего действительно не хватает, так это звука огня, который теряется под персонажами Шрека, время от времени блуждающими по экрану.
Подходит ли к Canon? В короткометражном спин-оффе Shrek the Halls установлено, что Рождество действительно существует во вселенной Шрека, и короткометражка о том, что это Рождественское бревно изначально было соединено с Рождественским шректакуляром Ослика , продолжает эту тенденцию. Мы сразу видим, где мы находимся — болотная гавань Шрека — и куда вписывается эта сцена: Шрек зажигает огонь прямо перед тем, как он и Фиона пригласят гостей на праздники. Мы — это во вселенной Шрека, тусуемся со Шреком и его семьей. Идеальные часы, если вы жаждете теплого семейного общения, но у вас есть только YouTube.
Сколько времени? Также немного меньше 30 минут, так что, как и в случае с Святочным журналом Шрека, вам нужно будет нажать повтор. Или играть их спина к спине!
Уровни рождественского настроения? «Бревно сноглетог» — это просто окно в фантастическое Рождество в мире Как приручить дракона , вплоть до украшений и веселой рождественской музыки, играющей на заднем плане. Как и в случае с видео Шрека, многие настоящие звуки огня теряются среди фоновой музыки и персонажей, блуждающих по экрану.
Подходит ли к Canon? Ага! Подобно тому, как Шрек был компаньоном «Рождественский шректакуляр осла», «Бревно сноглетог» является компаньоном «Как приручить дракона: Возвращение домой », премьера которого состоялась ранее в этом месяце на канале NBC. Действие короткометражного фильма происходит после событий финального фильма «Как приручить дракона », в котором исследуются последствия трилогии. У Иккинга и Астрид есть дети, а также у Беззубика и Дневной Фурии, и все они появляются здесь. Вполне вписывается в канон.
Сколько времени? Час!
Уровень рождественского настроения? Все начинается довольно скромно, но по ходу видео Соник бежит и получает все больше и больше декораций, пока весь очаг не становится невероятно праздничным. Там есть елка, чулки, подарки, список желаний для Санты и многое другое.
Подходит ли к Canon? У главных героев есть собственные чулки, и Соник, вероятно, из тех парней, которые очень рады праздникам.
В настоящее время их два: симпатичный анимированный и более реалистичный для Venom: Let There Be Carnage .
Сколько времени? Первая длится 10 часов, но кажется, что это всего лишь 10-секундный цикл анимированного Венома, который слегка подпрыгивает и пьет горячее какао. (Отказ от ответственности: мы не смотрели все 10 часов. Может быть, он делает что-то сложное и увлекательное за семь часов.)
Второй также длится 10 часов, при этом огни меняются и мигают через определенные промежутки времени.
Уровни рождественского настроения? Анимированный Веном сидит рядом с уютным камином под портретом Венома, одетого в причудливый шелковый жакет богатого человека.