
При свете табака
В лаборатории биотехнологов из Института биоорганической химии РАН выросли растения табака, которые осветили все вокруг мягким зеленым светом. На очереди — петуния, затем орхидеи или розы. Один из создателей растений, Илья Ямпольский, рассказал N + 1, как скоро живые светильники появятся в продаже, достаточно ли их света для чтения и почему это не просто игрушка, а новый инструмент для научных исследований.
Как это работает?
Сам эффект называется «биолюминесценция» — это нетепловое свечение в живой системе. Светятся светлячки, некоторые глубоководные рыбы, грибы, бактерии. Свечение происходит благодаря окислению молекул люциферинов ферментом люциферазой. Всего известно около 40 биолюминесцентных систем, включающих семь различных типов люцифераз.
Светящихся растений в природе не бывает. Сделать их искусственно пытались и раньше: например, десять лет назад группа под руководством Александра Кричевского встроила бактериальную люминесцентную систему в растения, но сделать их достаточно яркими не получилось: оказалось сложно совместить прокариотическую биохимическую цепочку с эукариотами.
В 2017 году наша группа описала люминесцентную систему грибов. Мы изучили синтез люциферина во вьетнамском светящемся грибе Neonotopanus nambi и выяснили, что грибной люциферин — это 3-гидроксигиспидин, который образуется из кофейной кислоты, обычного метаболита растений, поэтому химический цикл назвали «циклом кофейной кислоты». Через год мы определили все гены, отвечающие за этот процесс, что открыло возможность воспроизвести его в других организмах.
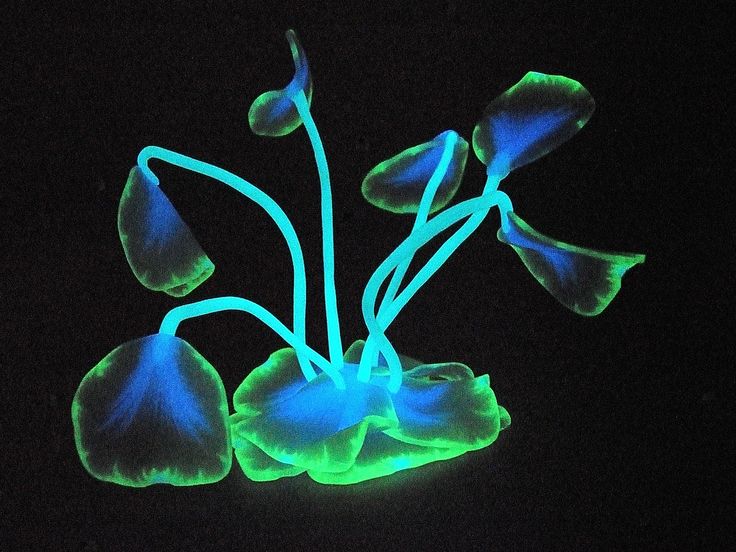
Примерно полгода назад наша группа, в которую вошли ученые из ИБХ РАН, компаний Planta и «Биотрон», впервые получили светящееся растение: вставили в геном табака Nicotiana tabacum гены гриба, которые кодируют ферменты синтеза грибного люциферина (гиспидина) из кофейной кислоты, ген люциферазы и фермент для превращения окисленного люциферина обратно в кофейную кислоту. Растения, которые мы получили, светятся в 10-100 раз ярче «бактериальных». Теперь результаты эксперимента, который поддерживало «Сколково» и РНФ, опубликованы в Nature Biotechnology.
Кофейная кислота есть во всех растениях, поэтому светиться, в принципе, можно заставить любое.
Как ярко?
Разные части растений светятся с разной яркостью. Например, листья выращенных нами растений светились с интенсивностью около 20 миллиардов фотонов в минуту на квадратный сантиметр, а цветы — порядка 30 миллиардов.
В темной комнате такие растения позволяют видеть предметы вокруг, стены. После того, как глаза привыкнут к темноте, при таком свете можно даже разобрать текст.
Постоянно или нет?
Растения светятся все время, непрерывно, но яркость может колебаться. Они начинают светиться ярче, если, например, положить рядом банановую кожуру. Она выделяет этилен, растительный гормон, который, с одной стороны, угнетает рост растений, а с другой — способствует созреванию плодов.
Интенсивность свечения колеблется в зависимости от времени суток, пик яркости приходится примерно на середину ночи. Молодые побеги светятся ярче, старые тусклее. Ярко светятся цветы. А если листу пора умирать, там включается какая-то биохимия, и он вспыхивает перед тем, как погибнуть окончательно.
Молодые побеги светятся ярче, старые тусклее. Ярко светятся цветы. А если листу пора умирать, там включается какая-то биохимия, и он вспыхивает перед тем, как погибнуть окончательно.
Очень ярко светятся корни, особенно точки их ветвления. Если отрезать побег, на этом месте начинает вырастать новый, и это место тоже светится очень ярко. Срезанная часть растения будет светиться до тех пор, пока она не засохнет.
А можно заставить светиться деревья? А животных?
Грибная люминесцентная система — единственная, которая может работать во всех эукариотах, не только в растениях. Поэтому принципиальной границы здесь нет, мы можем заставить светиться и другие организмы. С деревьями работать сложнее, поскольку у них значительно больше срок жизни. В случае животных придется модифицировать саму систему, но непреодолимых препятствий тут нет.
Зачем нужны такие растения?
Это совершенно новый инструмент для ученых, он позволит узнать о биохимических процессах в растениях множество вещей, которые раньше были нам совершенно недоступны.
В животных это делают с помощью GFP — зеленого флуоресцентного белка. Ген GFP пришивается к какому-то биологическому процессу, который вы хотите изучить, а дальше вы сидите и ждете, когда он «загорится» под действием ультрафиолета. Засветился — значит белок экспрессируется и помеченная вами система работает.
С растениями этот метод не работает. Растительная ткань очень плотно пигментирована — там и хлорофилл, и каротиноиды, они все флуоресцируют. Если растительную ткань поместить под микроскоп, вы увидите поток флюоресценции, на фоне которого увидеть полезный сигнал почти невозможно.
Поэтому использовались не флуоресцентные репортеры, а люминесцентные — те, которые не «отсвечивают» в ответ на излучение, а светятся сами. Наблюдать за ними, соответственно, надо в темноте. Но поскольку растения сами не вырабатывают люциферин, его надо физически вводить туда, где он должен сработать. Это неудобно и дорого.
А «кофейная» система не требует никаких дополнительных опрыскиваний, поливаний, она работает сама по себе, поскольку встроена в растительный метаболизм.
Потенциальных задач для такой системы очень много. Например, можно ее использовать для изучения реакций растений на стрессы — высокую температуру, высокую соленость, болезни, а потом с опорой на эти данные вывести устойчивые сорта. Десятки лабораторий со всего мира уже обратились к нам, запросили гены, семена — мы помогаем, отправляем им, например, готовые плазмиды.
Готового коммерческого продукта у нас нет. Но если к нам обратится компания, которая захочет выращивать такие растения на продажу, то они должны будут купить у нас лицензию.
Законно ли это в России?
Наши растения подпадают под регулирование, касающееся ГМ-организмов — но эксперименты с ГМ-растениями не запрещены, регулирование в основном касается выращивания модифицированных растений в открытом грунте.
Мы собираемся создать растения, которые смогут пройти все необходимые проверки и получить разрешение для продажи на рынке. Рассчитываем, что получение всех необходимых сертификатов и лицензий займет два-три года.
А купить такой росток можно?
Компания Planta не собирается продавать светящийся табак, мы сейчас работаем над получением светящихся разновидностей традиционных декоративных растений. Я не могу назвать их все, но скажу, что идет работа над розами, орхидеями, всего их около дюжины сортов. Первым светящимся растением на рынке будет, скорее всего, петуния, планируем начать продавать их уже через два года.
Мы хотим создать линейку растений и занять свои ниши во всех сегментах декоративных растений — от срезанных цветов до газонной травы и кустарников.
Текст подготовил Сергей Кузнецов
экономия на освещении улиц и стильный ландшафтный дизайн
Собака Баскервилей, своим светящимся в темноте оскалом доведшая до смерти сэра Чарльза Баскервиля и чуть было не уморившая Генри Баскервиля в романе Артура Конан Дойла, была обыкновенной собакой. Светилась она только благодаря злому умыслу и фосфору на морде. Однако живые организмы, самостоятельно светящиеся в темноте, действительно существуют.
Светилась она только благодаря злому умыслу и фосфору на морде. Однако живые организмы, самостоятельно светящиеся в темноте, действительно существуют.
Учёные пытаются создать светящиеся растения
Бактерии, медузы, моллюски, планктон, светлячки, скорпионы, грибы (в том числе и привычные опята). Сегодня учёным известно более восьми сот светящихся живых организмов. Большинство из них обитает в морях и океанах. Но вот представителей царства Флоры, обладающих способностью к биолюминесценции, учёные пока не обнаружили. Однако человек не привык ждать милостей от Природы: если она по какой-то причине «не додумалась» сделать светящиеся растения, «венец творения» готов сам взяться за это дело.
Ads by
«Bioglow» – компания, создавшая концепт светящегося растения
В природе нет светящихся растений, потому что растения не нуждаются в биолюминесценции. В микромире свечение – это побочное явление при пищеварении: нейтрализация активного кислорода ферментами бактерий при расщеплении глюкозы. Светлячки и маленькие кальмары-ватазении используют свет для привлечения партнёров, медузы – в качестве шоковой защиты от тех, кто пытается их съесть.
Светлячки и маленькие кальмары-ватазении используют свет для привлечения партнёров, медузы – в качестве шоковой защиты от тех, кто пытается их съесть.
Светящийся от планктона океан
Также есть охотники, привлекающие своих жертв свечением собственного тела. А некоторые виды глубоководных кораллов, по мнению учёных, способны слабый коротковолновый свет, проникающий в глубину, трансформировать в более яркие вспышки. Это явление используется как фитоподсветка для возможности фотосинтеза симбиотических колоний водорослей, живущих в коралловых зарослях.
Растениям светиться ни к чему. Поэтому потребовалось вмешательство генной инженерии, десятилетия работы и солидные капиталовложения. Хлоропласты растений – полуавтономные пластиды, существующие в симбиозе с растениями. Согласно гипотезе, когда-то они были самостоятельными, как и родственные хлоропластам цианобактерии, способные к свечению. Александр Кричевский (Сент-Луис, США) – специалист в двух областях: изучении явления биолюминесценции морских бактерий и микробиологии растений.
 У учёного возникла мысль об объединении двух хорошо знакомых ему дисциплин, что он и сделал, создав биотехнологическую компанию «Bioglow, Inc».
У учёного возникла мысль об объединении двух хорошо знакомых ему дисциплин, что он и сделал, создав биотехнологическую компанию «Bioglow, Inc».Starlight Avatar: воплощение звездного света
Компанией Александра Кричевского был создан концепт светящегося растения – «Starlight Avatar®».
Starlight Avatar — светящееся растение табака. Фото с сайта bioglowtech.com
Свечение Starlight Avatar (растения табака) основано на внедрении в геном растения части гена светлячка – молекулы люциферазы. Чтобы Starlight Avatar светился, необходим катализатор – реакция свечения происходит при окислении люциферазы под действием кислорода в присутствии фермента люциферина. Люциферин содержался в питательной среде, в которой выращивалось растение.
Свечение нового поколения генно-модифицированных растений (слева) в сравнении с Starlight Avatar (справа). Фото с сайта bioglowtech.com
В 2014 году на аукционе компанией Bioglow было продано двадцать экземпляров светящихся Starlight Avatar, растущих в специальных контейнерах. Пока этот свет очень слабый, но лаборатория Александра Кричевского работает над увеличением яркости.
Пока этот свет очень слабый, но лаборатория Александра Кричевского работает над увеличением яркости.
Пока – из области фантастики
В планах Bioglow – создание растений, которые не только смогут украсить ландшафт ночью, но и помогут сэкономить на уличном освещении. Но пока светящиеся растения – это из области фантастики. Starlight Avatar испускает свет, только если его поливать соответствующим раствором.
Российские учёные, работающие над исследованием биолюминесценции и созданием самостоятельно светящихся растений в лаборатории биомолекулярной спектроскопии Института биоорганической химии Российской Академии наук под руководством Ильи Ямпольского, считают, что пока ещё рано планировать клумбы на своих участках с учётом светящихся в темноте роз или пионов и выкорчёвывать живую изгородь, чтобы поменять её на светящуюся.
Они называют биолюминесценцию растений одним из самых амбициозных проектов: «Идеальный вариант, который пока не удался никому, включает в себя расшифровку всего пути биосинтеза люциферина, который может быть многоэтапным процессом с участием большого числа белков. Потом – встраивание в геном другого организма генов, кодирующих все эти белки и люциферазу. На данный момент расшифрован биосинтез только бактериального люциферина, однако эта система тяжело адаптируется к растениям и животным. И реализация такого подхода представляется маловероятной».
Потом – встраивание в геном другого организма генов, кодирующих все эти белки и люциферазу. На данный момент расшифрован биосинтез только бактериального люциферина, однако эта система тяжело адаптируется к растениям и животным. И реализация такого подхода представляется маловероятной».
Дорожка со светящимся гравием. Фото с сайта passages-ivm.com
Семян светящихся цветов пока ещё купить нельзя (если вы, конечно, не заплатили 40 $ за гипотетическую возможность получить семена генно-модифицированной резуховидки Таля в краудфандинговом проекте GLOWING PLANTS). Но не расстраивайтесь: зато можно приобрести искусственные светящиеся камни – для декора дорожек на своём участке, создания альпийских горок, видимых и в темноте, и даже для отделки фасада дома. Ну, или на крайний случай – хотя бы для декорирования аквариума.
Да будет свет | Природа Растения
Да будет свет
Скачать PDF
Скачать PDF
- Результаты исследований
- Опубликовано:
- Гийом Тена 1
Природные растения том 5 , страница 1103 (2019)Процитировать эту статью
4720 доступов
31 Альтметрический
Детали показателей
Субъекты
- Биолюминесцентная визуализация
- Биотехнология растений
- Репортерные гены
- Синтетическая биология
bioRxiv http://doi. org/ggbdcj (2019).
org/ggbdcj (2019).
bioRxiv http://doi.org/dc5x (2019).
Биолюминесцентные растения существовали всегда — но только в воображении любителей научной фантастики. В реальном мире различные виды насекомых, грибов, бактерий, морских животных, динофлагеллят и т. д. могут излучать свет сами по себе, но ни одно известное растение не проявляет аутогенной биолюминесценции. Ученые уже некоторое время пытаются исправить этот досадный эволюционный факт. Ранее были разработаны автономные светящиеся установки, не требующие добавления экзогенного субстрата с непредсказуемой диффузией. Эти редкие биоинженерные достижения, часто омраченные преувеличенными маркетинговыми заявлениями, глупыми заголовками и нереалистичными обещаниями заменить электрические лампочки, начались почти десять лет назад с транспластомного табака, преобразованного с помощью полной бактериальной люкс оперон.
Авторы и права: Nature Picture Library / Alamy Stock Photo
Теперь в двух препринтах, одновременно опубликованных на bioRxiv , две группы предлагают новый многообещающий подход, основанный на недавно открытом биохимическом пути люминесценции грибов. Трех генов достаточно, чтобы производить и перерабатывать люциферин из кофейной кислоты, вездесущего растительного метаболита, занимающего центральное место в фенилпропаноидном пути. Четвертый грибковый ген кодирует люциферазу, которая окисляет люциферин и производит видимые фотоны. Количество света, на порядок выше, чем в предыдущих попытках, достаточно, чтобы увидеть свечение невооруженным глазом или записать его всего за несколько секунд выдержки с помощью неспециализированных камер. Излучаемый свет зеленый, что сводит к минимуму помехи от фоторецепторов растений.
Трех генов достаточно, чтобы производить и перерабатывать люциферин из кофейной кислоты, вездесущего растительного метаболита, занимающего центральное место в фенилпропаноидном пути. Четвертый грибковый ген кодирует люциферазу, которая окисляет люциферин и производит видимые фотоны. Количество света, на порядок выше, чем в предыдущих попытках, достаточно, чтобы увидеть свечение невооруженным глазом или записать его всего за несколько секунд выдержки с помощью неспециализированных камер. Излучаемый свет зеленый, что сводит к минимуму помехи от фоторецепторов растений.
Одна команда из России получила трансгенные растения табака со сверхэкспрессией всех четырех генов и изучила динамику и распределение люминесценции, отражающей изменения метаболизма фенилпропаноидов, и его реакцию на внешние сигналы. Другая группа из США сделала следующий логический шаг и использовала тот же набор грибковых инструментов в качестве автолюминесцентного количественного репортера для изучения экспрессии специфического гена. Переходные тесты показывают, что этот подход работает на различных видах растений. Затем ген люциферазы управляется циркадианными и ABA-регулируемыми промоторами. Динамика люминесценции следует ожидаемым закономерностям, обнаруживаемым с помощью дешевой установки.
Переходные тесты показывают, что этот подход работает на различных видах растений. Затем ген люциферазы управляется циркадианными и ABA-регулируемыми промоторами. Динамика люминесценции следует ожидаемым закономерностям, обнаруживаемым с помощью дешевой установки.
Новый подход, описанный в этих двух исследованиях, все еще находится на стадии проверки концепции. Необходима оптимизация, особенно переработка окисленного люциферина, чтобы избежать истощения субстрата, необходимого как для излучения света, так и для метаболизма растений. Но нет никаких сомнений в том, что автономные биолюминесцентные растения в ближайшем будущем станут плюсом для исследований, требующих легко отслеживаемых биосенсоров и генных маркеров. Сейчас происходит научная фантастика.
Информация об авторе
Авторы и принадлежности
Природные растения https://www.nature.com/nlants
Гиома Тена
Авторы
- PubMed Google Scholar
Автор, ответственный за корреспонденцию
Гийом Тена.
Права и разрешения
Перепечатка и разрешения
Об этой статье
Растения с генетически закодированной автолюминесценцией
Доступность данных
Наборы данных, полученные или проанализированные в текущем исследовании, можно получить у соответствующих авторов по обоснованному запросу. Необработанные изображения люминесцентных цветов, снятые на камеру Sony Alpha ILCE-7M3 и IVIS Spectrum CT, доступны на сайте Figshare (https://doi.org/10.6084/m9.figshare.11353601). Последовательности плазмид доступны в Genbank под следующими инвентарными номерами: pHpaB-C1, MT233533; pHpaC-C1, МТ233534; pnnCPH-C1, MT233535; pnnh4H-C1, МТ233536; pnnHispS-C1, MT233537; pnnLuz-C1, MT233538; pnpgA-C1, MT233539; pRCTAL-C1, MT233540; рХ037, МТ233541. Результаты секвенирования Sanger и Illumina доступны в качестве дополнительных данных.
Change history
04 June 2020
A Correction to this paper has been published: https://doi.
 org/10.1038/s41587-020-0578-0
org/10.1038/s41587-020-0578-0
 org/10.1038/s41587-020-0578-0
org/10.1038/s41587-020-0578-0References
Krichevsky, А., Мейерс, Б., Вайнштейн, А., Малига, П. и Цитовски, В. PLoS ONE 5 , e15461 (2010).
Артикул Google Scholar
Котлобай А.А. и др. Проц. Натл акад. науч. США 115 , 12728–12732 (2018).
Артикул КАС Google Scholar
Yan, Y. & Lin, Y. Биосинтез кофейной кислоты и производных кофейной кислоты рекомбинантными микроорганизмами. Патент США 8809028B2 (2012 г.).
Кавамата, С. и др. Физиол клеток растений. 38 , 792–803 (1997).
Артикул КАС Google Scholar
Гакерель, Э., Гулати, Дж. и Болдуин, И.
Т. Plant J. 79 , 679–692 (2014).Артикул КАС Google Scholar
Тойота, М. и др. Наука 361 , 1112–1115 (2018).
Артикул КАС Google Scholar
Singh, S.K. et al. науч. Респ. 5 , 18148 (2015).
Артикул КАС Google Scholar
Ли, Л. и др. науч. Респ. 6 , 37976 (2016).
Артикул КАС Google Scholar
Ли, В. и др. науч. Респ. 7 , 12126 (2017).
Артикул Google Scholar
Ву, Х. Р., Ким, Х. Дж., Нам, Х. Г. и Лим, П. О. J. Cell Sci. 126 , 4823–4833 (2013).
КАС пабмед Google Scholar
Pauwels, L. et al. Проц. Натл акад. науч. США 105 , 1380–1385 (2008 г.).
Артикул КАС Google Scholar
Bernards, M. A. & Båstrup-Spohr, L. Индуцированная устойчивость растений к травоядным (Springer, 2008).
Сингх, Р., Растоги, С. и Двиведи, ООН Compr. Преподобный Food Sci. Пищевая безопасность 9 , 398–416 (2010).
Артикул КАС Google Scholar
Weber, E., Engler, C., Gruetzner, R., Werner, S. & Marillonnet, S. PLoS ONE 6 , e16765 (2011).
Артикул КАС Google Scholar
Айверсон С.В., Хэддок Т.Л., Бил Дж. и Денсмор Д.
М. ACS Synth. биол. 5 , 99–103 (2016).Артикул КАС Google Scholar
Лазо, Г. Р., Штейн, П. А. и Людвиг, Р. А. Биотехнология 9 , 963–967 (1991).
Артикул КАС Google Scholar
Роджерс С.О. и Бендич А.Дж. Руководство по молекулярной биологии растений (Springer, 1994).
 Т. Plant J. 79 , 679–692 (2014).
Т. Plant J. 79 , 679–692 (2014).
 М. ACS Synth. биол. 5 , 99–103 (2016).
М. ACS Synth. биол. 5 , 99–103 (2016).Скачать ссылки
Благодарности
Это исследование было разработано, проведено и профинансировано ООО «Планта». Мы благодарим К. Вуда за помощь в разработке рукописи. Планта выражает благодарность Инновационному центру «Сколково» за поддержку. Мы благодарим Д. Болотина и Милабораторию (milaboratory.com) за доступ к вычислительной инфраструктуре и инфраструктуре хранения. Благодарим С. Шахова за предоставленную фототехнику. Группа синтетической биологии финансируется Лондонским институтом медицинских наук MRC (UKRI MC-A658-5QEA0, KSS). К.С.С. поддерживается исследовательской стипендией Имперского колледжа. Эксперименты частично проведены на оборудовании, предоставленном БХ ИБХ РАН (ЦКП ИБХ; поддержан грантом Минобрнауки России RFMEFI62117X0018). Ф.А.К. лаборатория поддерживается соглашением о гранте ERC 771209—ЧарФЛ. Этот проект получил финансирование от исследовательской и инновационной программы Horizon 2020 Европейского Союза в рамках Соглашения о гранте Марии Склодовской-Кюри 665385. K.S.S. признает поддержку президентским грантом 075-15-2019-411. Дизайн и сборка некоторых плазмид поддержаны грантом РНФ 19-74-10102. Эксперименты по визуализации частично поддержаны грантом РНФ 17-14-01169р. ЖХ-МС/МС анализы экстрактов были поддержаны грантом РНФ 16-14-00052р. Дизайн и сборка плазмид частично поддержаны грантом 075-15-2019-1789 от Министерства науки и высшего образования Российской Федерации выделен Центру точного редактирования генома и генетических технологий для биомедицины. Авторы выражают признательность Центру геномики Сколковского института науки и технологий за работу по секвенированию и биоинформационному анализу.
К.С.С. поддерживается исследовательской стипендией Имперского колледжа. Эксперименты частично проведены на оборудовании, предоставленном БХ ИБХ РАН (ЦКП ИБХ; поддержан грантом Минобрнауки России RFMEFI62117X0018). Ф.А.К. лаборатория поддерживается соглашением о гранте ERC 771209—ЧарФЛ. Этот проект получил финансирование от исследовательской и инновационной программы Horizon 2020 Европейского Союза в рамках Соглашения о гранте Марии Склодовской-Кюри 665385. K.S.S. признает поддержку президентским грантом 075-15-2019-411. Дизайн и сборка некоторых плазмид поддержаны грантом РНФ 19-74-10102. Эксперименты по визуализации частично поддержаны грантом РНФ 17-14-01169р. ЖХ-МС/МС анализы экстрактов были поддержаны грантом РНФ 16-14-00052р. Дизайн и сборка плазмид частично поддержаны грантом 075-15-2019-1789 от Министерства науки и высшего образования Российской Федерации выделен Центру точного редактирования генома и генетических технологий для биомедицины. Авторы выражают признательность Центру геномики Сколковского института науки и технологий за работу по секвенированию и биоинформационному анализу.
Информация об авторе
Примечания автора
Авторы внесли одинаковый вклад: Татьяна Митюшкина, Александр Сергеевич Мишин, Луиза Гонсалес Сомермейер, Надежда М. Маркина, Илья В. Ямпольский, Карен С. Саркисян.
Авторы и аффилированные лица
ООО «Планта», Москва, Россия
Митюшкина Татьяна Владимировна, Мишин Александр Сергеевич, Маркина Надежда Михайловна, Чепурных Татьяна Владимировна, Чепурных Татьяна Александровна, Каратаева Е.С., Палкина Е. Ксения. Шахова, Лилия И. Фахранурова, Софья В. Чекова, Александра Сергеевна Царькова, Олеся А. Мельник, Александр Сергеевич Пушин, Илья Владимирович Ямпольский и Карен Сергеевич Саркисян
Институт биоорганической химии им. наук, Москва, Россия
Митюшкина Татьяна, Мишин Александр Сергеевич, Маркина Надежда Михайловна, Чепурных Татьяна Владимировна, Гугля Елена Борисовна, Каратаева Татьяна Александровна, Палкина Ксения Александровна, Шахова Екатерина Сергеевна, Фахранурова Лилия Ивановна, Царькова Александра Сергеевна, Дмитрий Шлыков, Олеся А.
Мельник, Виктория О. Шипунова, Сергей М. Деев, Андрей И. Бубырев, Александр Сергеевич Пушин, Сергей В. Долгов, Илья Владимирович Ямпольский и Карен Сергеевич СаркисянИнститут науки и Technology Austria, Клостернойбург, Австрия
Луиза Гонсалес Сомермейер и Фёдор А. Кондрашов
Российский национальный исследовательский медицинский университет им. Н. И. Пирогова, Москва, Россия
Елена Б. Гугля, Ярослав В. Голубев, Вадим В. Негребецкий и Илья В. Ямпольский0 0 Институт Биофизика, Красноярский научный центр СО РАН, Красноярск, Россия
Царькова Александра Сергеевна
ООО «Айвок», Зеленоград, Москва, Россия
Сергей А. Долгушин и Павел В. Шалаев
Ботанический сад Московского государственного университета им. М.В. Ломоносова, Москва, Россия
Владимир В. Чуб
Группа синтетической биологии, MRC Лондонский институт медицинских наук, Лондон, Великобритания
Саркисян Карен С.
Институт клинических наук медицинского факультета и Центр синтетической биологии Имперского колледжа, Имперский колледж Лондона, Лондон, Великобритания
Саркисян Карен С.
 Мельник, Виктория О. Шипунова, Сергей М. Деев, Андрей И. Бубырев, Александр Сергеевич Пушин, Сергей В. Долгов, Илья Владимирович Ямпольский и Карен Сергеевич Саркисян
Мельник, Виктория О. Шипунова, Сергей М. Деев, Андрей И. Бубырев, Александр Сергеевич Пушин, Сергей В. Долгов, Илья Владимирович Ямпольский и Карен Сергеевич Саркисян
Авторы
- Татьяна Митюшкина
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Мишин Александр Сергеевич
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Louisa Gonzalez Somermeyer
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Маркина Надежда Михайловна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Татьяна Владимировна Чепурных
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Гугля Елена Борисовна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Каратаева Татьяна Анатольевна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ксения А. Палкина
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Екатерина Сергеевна Шахова
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Фахранурова Лилия Ивановна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- София В. Чекова
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Александра Сергеевна Царькова
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Голубев Ярослав Васильевич
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Негребецкий Вадим В.
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Сергей А. Долгушин
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Шалаев Павел Васильевич
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Дмитрий Шлыков
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Мельник Олеся Александровна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Шипунова Виктория Олеговна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Сергей М. Деев
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Андрей Иванович Бубырев
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Пушин Александр Сергеевич
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Владимир В. Чуб
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Сергей В. Долгов
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Кондрашов Ф.А.
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ямпольский Илья Владимирович
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Саркисян Карен С.
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
 Палкина
Палкина
 Чуб
ЧубВзносы
T.M., A.S.M., L.G.S., T.V.C., E.B.G., T.A.K., N.M.M., S.V.C., A.S.T., L.I.F., K.A.P., E.S.S. и К.С.С. проводили эксперименты. T.M., A.S.M., L.G.S., T.V.C., E.B.G., T.A.K., N.M.M., S.V.C., A. S.T., L.I.F., K.A.P., E.S.S., Y.V.G., V.V.N., S.A.D., A.V. и К.С.С. провел анализ данных. КАК М. разработал установку для визуализации, спланировал и провел эксперименты, проанализировал данные и написал статью. И.В.Ю. и К.С.С. предложил и руководил исследованием, планировал эксперименты и написал статью. Все авторы рассмотрели и прокомментировали проект статьи.
S.T., L.I.F., K.A.P., E.S.S., Y.V.G., V.V.N., S.A.D., A.V. и К.С.С. провел анализ данных. КАК М. разработал установку для визуализации, спланировал и провел эксперименты, проанализировал данные и написал статью. И.В.Ю. и К.С.С. предложил и руководил исследованием, планировал эксперименты и написал статью. Все авторы рассмотрели и прокомментировали проект статьи.
Авторы переписки
Переписка с Илья В. Ямпольский или Карен С. Саркисян.
Декларации этики
Конкурирующие интересы
Работа выполнена при поддержке ООО «Планта». И.В.Ю. и К.С.С. являются акционерами и сотрудниками Planta. Планта подала патентные заявки, связанные с использованием компонентов грибковой биолюминесцентной системы и разработкой светящихся трансгенных организмов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Интегрированная дополнительная информация
Дополнительная информация Рисунок 1. Инженерия автономной люминесценции в клетках млекопитающих.
и . Наложение фонового света (черно-белое) и люминесцентных (псевдоцветных) изображений многолуночного планшета с клетками HEK293T, котрансфицированными плазмидами, кодирующими гены пути биолюминесценции грибов и биосинтеза кофейной кислоты: rctal, hpab, hpac, nnhisps, npga, nnh4h, nncph, nnluz . Репрезентативное люминесцентное изображение трех независимых экспериментов. б . Световая эмиссия автолюминесцентных клеток в стандартной среде МЕМ с 20 мМ HEPES и в той же среде с добавлением кофейной кислоты. Точки данных были собраны в трех независимых экспериментах.
Дополнительный рисунок 2.
Структура плазмиды, используемой для создания линий растений с самоподдерживающейся люминесценцией.
Дополнительный рисунок 3. Фенотип светящегося и дикого типа
Nicotiana tabacum растения.
Горшечные светящиеся растения (линия NT001) и растения дикого типа (линия NT000) одного возраста при рассеянном свете ( a , b ) и в темноте ( c ). Четырехнедельные растения на панели и являются репрезентативными для 100 растений дикого типа и 100 светящихся растений, выращенных in vitro в четырех отдельных экспериментах. д . Сравнение содержания хлорофилла а, хлорофилла b и каротиноидов в листьях, размерах листьев и высоте растений 8-недельных растений трансгенной линии NT001 и линии дикого типа NT000. Рамки простираются от нижнего к верхнему квартилю данных, оранжевая линия представляет собой медиану. Усы представляют полный диапазон данных. Точные размеры выборки указаны над каждым прямоугольным графиком, статистика (U) и p-значения двунаправленного U-критерия Манна-Уитни отмечены в нижней правой части каждого графика.
Дополнительный рисунок 4. Биолюминесцентные растения
Nicotiana tabacum и Nicotiana benthamiana .
a и b — фотографии растений в горшках при естественном освещении и в темноте, соответственно, снятые на камеру Sony Alpha ILCE-7M3. Листья, отмеченные розовыми треугольниками на и , а также контрольные листья растений дикого типа затем срезали и визуализировали в IVIS Spectrum CT при естественном освещении и в темноте отдельно для Nicotiana tabacum 9.0024 ( c , d ) и Nicotiana benthamiana ( e , f ). Черную бумажную коробку вокруг контрольных листьев, видимых на изображениях c и e , использовали для предотвращения освещения листьев контрольных растений светом, излучаемым трансгенными растениями. Все изображения соответствуют одному эксперименту, выполненному на трех светящихся и одном диком типе Nicotiana tabacum , а также на трех светящихся и одном диком типе Nicotiana benthamiana 9.0024 растения.
Дополнительный рисунок 5. Цветы, собранные из линий дикого типа и биолюминесцентных
Nicotiana tabacum и Nicotiana benthamiana , изображение на IVIS Spectrum CT.
Nicotiana tabacum ( a , b ) и Nicotiana benthamiana ( c , d ). a и c — фотографии при естественном освещении, b и d — фотографии в темноте. Черная бумажная коробка вокруг контрольных цветов видна на изображениях 9.0131 а и с использовали для предотвращения освещения цветков контрольных растений светом, излучаемым трансгенными растениями. Все изображенные цветы были случайным образом собраны с разных растений в одном эксперименте по визуализации (6 светящихся и 3 растения дикого типа Nicotiana tabacum , 6 светящихся и 4 растения дикого типа Nicotiana benthamiana ).
Дополнительный рисунок 6. Количественное сравнение светового излучения цветков
Nicotiana tabacum (правая сторона каждой фотографии) и Nicotiana benthamiana (слева на каждой фотографии). a и b — это изображения при естественном освещении и в темноте, полученные с помощью IVIS Spectrum CT. c и d — это изображения одних и тех же цветов при естественном освещении и в темноте, снятые на камеру Sony Alpha ILCE-7M3. На c и d цветы были изображены вместе с калиброванным источником света XLS-4, который излучает 1,6*10 9 фотонов/сек при 525 нм. Этот откалиброванный источник света был отдельно отображен на IVIS Spectrum CT, см. Figshare (doi: 10,6084/m9)..figshare.11871888) для необработанных файлов. Все цветы были собраны с разных растений в одном эксперименте по визуализации (14 светящихся растений Nicotiana tabacum и 13 светящихся растений Nicotiana benthamiana ).
c и d — это изображения одних и тех же цветов при естественном освещении и в темноте, снятые на камеру Sony Alpha ILCE-7M3. На c и d цветы были изображены вместе с калиброванным источником света XLS-4, который излучает 1,6*10 9 фотонов/сек при 525 нм. Этот откалиброванный источник света был отдельно отображен на IVIS Spectrum CT, см. Figshare (doi: 10,6084/m9)..figshare.11871888) для необработанных файлов. Все цветы были собраны с разных растений в одном эксперименте по визуализации (14 светящихся растений Nicotiana tabacum и 13 светящихся растений Nicotiana benthamiana ).
Дополнительный рисунок 7. Выбор области интереса для анализа светового излучения цветов, используемого для получения значений яркости в дополнительной таблице 1 (обратите внимание, что числа на изображении представляют собой общий поток фотонов в секунду для каждой области интереса, а не среднюю яркость) .
Анализ нормализованного общего потока в обозначенных ROI доступен на Figshare (doi: 10.![]() 6084/m9.figshare.11871888). Цветы, показанные на этой фигуре, такие же, как и на дополнительной фигуре 6 .
6084/m9.figshare.11871888). Цветы, показанные на этой фигуре, такие же, как и на дополнительной фигуре 6 .
Дополнительный рисунок 8. Фотографии светящегося растения
Nicotiana benthamiana , сделанные на смартфон при естественном освещении и в темноте с 30-секундной выдержкой.( A ) Снято на смартфон при естественном освещении. ( Б ) Снято в темноте с выдержкой 30 секунд. Изображения являются результатом одного эксперимента.
Дополнительный рисунок 9. Инфильтрация листьев
Nicotiana benthamiana растворами предшественников гиспидина. А . Типичная фотография трансгенных биолюминесцентных листьев Nicotiana benthamiana , которым вводили раствор индивидуальных предшественников гиспидина. Изображения являются репрезентативными для четырех экспериментов, повторенных независимо друг от друга с аналогичными результатами Б, С . Инъекции гиспидина и люциферина в листья биолюминесцентных и диких растений N. benthamiana . Эксперимент был повторен дважды независимо с аналогичными результатами. Д . Блочные диаграммы, показывающие реакцию люминесценции в местах инъекций смесей предшественников гиспидина различного состава. Показаны точки данных. Изменение люминесценции – это абсолютное изменение сигнала (ΔL = люминесценция после инъекции – начальная люминесценция). Зачеркнутый текст подчеркивает отсутствие соединения в смеси. Примечательно, что только смеси, содержащие кофейную кислоту, вызывали усиление люминесценции. Поле простирается от нижнего к верхнему квартилю значений данных, оранжевая линия представляет собой медиану. Усы представляют полный диапазон данных. Группы, содержащие несколько реагентов (H-критерий Крускала-Уоллиса, H-статистика = 48,59)., p=7,11e-10) и отдельные потенциальные реагенты (H-критерий Крускала-Уоллиса, H-статистика: 68,65, p=8,3e-15) анализировали отдельно. P-значения апостериорных парных U-тестов Манна-Уитни, подчеркивающих влияние кофейной кислоты на люминесцентный отклик, указаны под скобками между диаграммами E .
benthamiana . Эксперимент был повторен дважды независимо с аналогичными результатами. Д . Блочные диаграммы, показывающие реакцию люминесценции в местах инъекций смесей предшественников гиспидина различного состава. Показаны точки данных. Изменение люминесценции – это абсолютное изменение сигнала (ΔL = люминесценция после инъекции – начальная люминесценция). Зачеркнутый текст подчеркивает отсутствие соединения в смеси. Примечательно, что только смеси, содержащие кофейную кислоту, вызывали усиление люминесценции. Поле простирается от нижнего к верхнему квартилю значений данных, оранжевая линия представляет собой медиану. Усы представляют полный диапазон данных. Группы, содержащие несколько реагентов (H-критерий Крускала-Уоллиса, H-статистика = 48,59)., p=7,11e-10) и отдельные потенциальные реагенты (H-критерий Крускала-Уоллиса, H-статистика: 68,65, p=8,3e-15) анализировали отдельно. P-значения апостериорных парных U-тестов Манна-Уитни, подчеркивающих влияние кофейной кислоты на люминесцентный отклик, указаны под скобками между диаграммами E . Влияние трех последовательных инъекций предшественников люциферина на интенсивность свечения листьев N. benthamiana .
Влияние трех последовательных инъекций предшественников люциферина на интенсивность свечения листьев N. benthamiana .
Доп. рисунок 10.
Фото в разрезе цветов из светящегося Nicotiana tabacum растение при рассеянном свете ( A ) и в темноте ( B ). Изображения двух цветов разных растений, изображенные на рисунке, являются репрезентативными для шести срезанных цветов трех растений.
Дополнительный рисунок 11. Динамика биолюминесценции после повреждений листьев.
А . Репрезентативное изображение листа до травмы. Б . Фото места разреза крупным планом. С . Динамика биолюминесценции в месте разреза после травмы (время экспозиции — 5 секунд). Масштабная линейка соответствует 1 см. Видео версия панели C доступен как дополнительное видео 7. Эксперимент с повреждением листьев был повторен трижды с аналогичными результатами.
Дополнительный рисунок 12. Устойчивое увеличение биолюминесценции боковых побегов, вызванное обрезкой.

А . Фото при рассеянном свете. Боковые побеги отмечены желтыми стрелками. Срез выделен желтой пунктирной линией на увеличенном фото (внизу). Б . Фото сделано в темноте. Изображения являются репрезентативными для двух экспериментов по визуализации, проведенных на шести растениях с аналогичными результатами.
Дополнительный рисунок 13. Динамика люминесценции у целых растений.
и . Фотография трансгенного растения Nicotiana tabacum , опрысканного 5 мМ раствором метилжасмоната или буфером (контроль) до и после обработки. Два изображенных растения являются репрезентативными для шести растений, проанализированных в трех независимых экспериментах по визуализации. б . Фотография трансгенного растения Nicotiana tabacum , инкубированного в закрытом флаконе со спелой кожурой банана. Два изображенных растения являются репрезентативными для четырех растений, проанализированных в двух независимых экспериментах по визуализации. с . Суточные колебания люминесценции. Фотографии трех растений снимались постоянно в течение десяти дней в условиях нормального освещения (17,5 ч в день; дни 1-3 и 8-10) или в постоянной темноте (дни 4-7). Каждое изображение растения 1 было сжато в одну вертикальную линию пикселей для создания кимограммы. Средняя яркость растений 1-3 отображается на графике. График в дни 1-3 репрезентативен для девяти растений в трех независимых экспериментах. Полный график (дни 1-10) показывает свечение трех растений в одном эксперименте. Псевдоцвет используется для визуальной ясности.
с . Суточные колебания люминесценции. Фотографии трех растений снимались постоянно в течение десяти дней в условиях нормального освещения (17,5 ч в день; дни 1-3 и 8-10) или в постоянной темноте (дни 4-7). Каждое изображение растения 1 было сжато в одну вертикальную линию пикселей для создания кимограммы. Средняя яркость растений 1-3 отображается на графике. График в дни 1-3 репрезентативен для девяти растений в трех независимых экспериментах. Полный график (дни 1-10) показывает свечение трех растений в одном эксперименте. Псевдоцвет используется для визуальной ясности.
Дополнительная фигура 14. Саузерн-блот ДНК, выделенной из различных светящихся трансгенных линий табака, с отжигом зонда до гена
nnluz . и . Саузерн-блот ДНК, выделенной из различных светящихся трансгенных линий табака, с отжигом зонда до гена nnluz . Результаты блота свидетельствуют о наличии двух копий гена nnluz в геноме линии NT001 (используемой для всех экспериментов N. tabacum в данном исследовании). Саузерн-блоттинг получен из одного эксперимента, проведенного на образцах ДНК, выделенных из пяти линий трансгенных растений и одной контрольной линии. б . Необработанное изображение полного сканирования того же пятна.
tabacum в данном исследовании). Саузерн-блоттинг получен из одного эксперимента, проведенного на образцах ДНК, выделенных из пяти линий трансгенных растений и одной контрольной линии. б . Необработанное изображение полного сканирования того же пятна.
Дополнительный рисунок 15. Закономерности динамики биолюминесценции в листьях молодых светящихся растений.
Наложение фотографий при естественном освещении и в темноте показано на A . Динамика люминесценции от двух растений, отмеченных квадратом на A, отображается на B . Видеоверсия панели B доступна в качестве дополнительного видео 9. Изображения на панели B представляют шесть растений в этом эксперименте, и был проведен еще один цейтраферный эксперимент с тремя растениями с аналогичными результатами.
Дополнительный рисунок 16. Фотографии поперечного сечения стебля светящегося растения
Nicotiana tabacum при естественном освещении и в темноте. ( A ) При внешнем освещении. ( B ) в темноте. Мощное свечение принадлежит клеткам паренхимы сердцевины побега (в центре стебля). Более слабая биолюминесценция наблюдается у клеток сердцевинной паренхимы (периферия стебля). Максимальная интенсивность света соответствует области пазушной почки — новообразованной меристеме — и окружающим паренхиматозным клеткам. Полностью одревесневшие ткани, ксилема и флоэма, с камбием между ними, лишены люминесценции (заметна на правой стороне стебля в виде темной полосы). Изображения представляют два эксперимента с аналогичными результатами.
( B ) в темноте. Мощное свечение принадлежит клеткам паренхимы сердцевины побега (в центре стебля). Более слабая биолюминесценция наблюдается у клеток сердцевинной паренхимы (периферия стебля). Максимальная интенсивность света соответствует области пазушной почки — новообразованной меристеме — и окружающим паренхиматозным клеткам. Полностью одревесневшие ткани, ксилема и флоэма, с камбием между ними, лишены люминесценции (заметна на правой стороне стебля в виде темной полосы). Изображения представляют два эксперимента с аналогичными результатами.
Дополнительная фигура 17. Экспрессия
nnluz в N. tabacum в течение дня, измеренная с помощью количественной ПЦР на 9 образцах кДНК, полученных из растений, выращенных в условиях естественного освещения. Все точки данных, включая технические повторы, показаны точками, каждый цвет соответствует разному биологическому образцу. Синие точки представляют собой средние значения Ct в каждый момент времени. Эксперимент проводился один раз.
Эксперимент проводился один раз.
Дополнительная информация
Дополнительная информация
Дополнительные рис. 1–17, таблицы 1–3 и примечания 1–7.
Резюме отчета
Доп. видео 1
Инъекция гиспидина в листья автономно люминесцентного N. tabacum. Покадровая люминесцентная визуализация после инъекции гиспидина (в центральную область верхней части пластинки) и кофейной кислоты (в апикальную часть) листьев автономно люминесцентного N. tabacum . Интенсивность люминесценции показана псевдоцветом. Видео представляет два эксперимента по визуализации, проведенных на шести растениях.
Дополнительное видео 2
Проращивание семян T1. Покадровая визуализация трансгенных семян N. tabacum T1, прорастающих в прозрачном агаре. Показано наложение фотографий в рассеянном свете (серый) и свечении (псевдоцвет). Видео пяти прорастающих семян является репрезентативным для двух экспериментов по визуализации с аналогичными результатами (всего 32 проростка).
Дополнительное видео 3
Регенерация корней. Покадровая люминесцентная визуализация регенерирующих корней срезанных побегов трансгенных N. tabacum растения (фото каждые 5 мин). Интенсивность люминесценции показана псевдоцветом. Видео корней двух отдельных растений является репрезентативным для трех независимых экспериментов по визуализации шести растений.
Дополнительное видео 4
Микроскопия корня. Покадровая микроскопия люминесцентного изображения регенерирующих корней срезанных побегов трансгенных растений N. tabacum (фотографии каждые 5 мин). Интенсивность люминесценции показана псевдоцветом. Видео показывает корни отдельного растения и является репрезентативным для двух независимых экспериментов по визуализации с большим увеличением с аналогичными результатами.
Дополнительное видео 5
Длительная съемка всей жизни растений. Покадровая визуализация трансгенных растений N. tabacum от вегетации до цветения. Интенсивность люминесценции показана псевдоцветом. Желтые звездочки обозначают дневное время. Этот длительный интервал времени является результатом одного эксперимента
Интенсивность люминесценции показана псевдоцветом. Желтые звездочки обозначают дневное время. Этот длительный интервал времени является результатом одного эксперимента
Дополнительное видео 6
Интервальная съемка цветов. Покадровое свечение цветков автономно люминесцентного N. tabacum . Интенсивность люминесценции показана псевдоцветом. Изображенные цветы одного растения являются репрезентативными для экспериментов по визуализации с аналогичными результатами.
Дополнительное видео 7
Повреждения листьев у N. tabacum. Покадровая люминесцентная визуализация после повреждения листовой пластинки автономно люминесцентного N. tabacum . Интенсивность люминесценции показана псевдоцветом. Опыт с повреждением листьев был повторен трижды с аналогичными результатами.
Дополнительное видео 8
Динамическое свечение после обрезки. Покадровая люминесцентная визуализация индуцированных обрезкой боковых побегов автономно люминесцентного Н.